Effects of laminectomy and TRPV1 modulation on calcium signaling and behavioral outcomes in DRG neurons
1. A
B
30μm
30μm
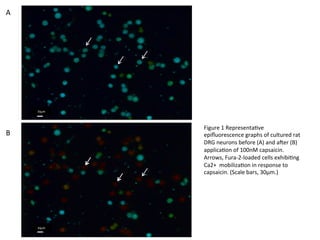
Figure
1
Representa5ve
epifluorescence
graphs
of
cultured
rat
DRG
neurons
before
(A)
and
aCer
(B)
applica5on
of
100nM
capsaicin.
Arrows,
Fura-‐2-‐loaded
cells
exhibi5ng
Ca2+
mobiliza5on
in
response
to
capsaicin.
(Scale
bars,
30μm.)
3. Figure
3
On
POD1,
potassium
chloride
evoked
calcium
transients
in
rat
DRG
neurons
as
measured
by
the
ra5o
difference
between
the
peak
ra5o
and
baseline
ra5o
(∆Ra5o
340/380).
Data
are
presented
as
mean±SEM
of
∆Ra5o
340/380.
***p<0.001,
**p<0.01,
n=50-‐80
cells
from
4-‐6
rats
each
group.
Figure
4
On
POD1,
dura5on
(seconds)
of
recovery
to
80%
of
the
baseline
of
K+
evoked
Ca2+
transients
in
rats
DRG
neurons.
Data
are
presented
as
mean±SEM
of
the
5me
dura5on
to
80%
recovery
of
baseline
of
∆Ra5o
340/380.
***p<0.001,
**p<0.01,
n=
50-‐80
cells
from
4-‐6
rats
each
group.
4. Figure
5
On
POD1,
capsaicin
evoked
calcium
transients
in
rat
DRG
neurons
as
measured
by
the
∆Ra5o
340/380.
Data
are
presented
as
mean±SEM
of
∆Ra5o
340/380.
***p<0.001,
*p<0.05,
n=45-‐70
cells
from
4-‐6
rats
each
group.
Figure
6
On
POD1,
dura5on
(seconds)
of
recovery
to
80%
of
the
baseline
of
capsaicin
evoked
Ca2+
transients
in
rats
DRG
neurons.
Data
are
presented
as
mean±SEM
of
the
5me
dura5on
to
80%
recovery
of
baseline
of
∆Ra5o
340/380.
***p<0.001,
*p<0.05,
n=
45-‐70
cells
from
4-‐6
rats
each
group.
Figure
7
On
POD1,
percentage
of
capsaicin
responsive
DRG
neurons.
Data
are
presented
as
mean±SEM
of
the
percentage
of
neurons
responsive
to
capsaicin
in
the
whole
DRG.
***p<0.001,
*p<0.05,
n=
45-‐70
cells
from
4-‐6
rats
each
group.
5. Figure
8
On
POD7,
potassium
chloride
evoked
calcium
transients
in
rat
DRG
neurons
as
measured
by
the
ra5o
difference
between
the
peak
ra5o
and
baseline
ra5o
(∆Ra5o
340/380).
Data
are
presented
as
mean±SEM
of
∆Ra5o
340/380.
**p<0.01,
n=45-‐70
cells
from
4-‐6
rats
each
group.
Figure
9
On
POD7,
dura5on
(seconds)
of
recovery
to
80%
of
the
baseline
of
K+
evoked
Ca2+
transients
in
rats
DRG
neurons.
Data
are
presented
as
mean±SEM
of
the
5me
dura5on
to
80%
recovery
of
baseline
of
∆Ra5o
340/380.
**p<0.01,
n=
45-‐70
cells
from
4-‐6
rats
each
group.
la
m
in
e
c
to
m
y
s
h
a
m
s
u
rg
e
ry
0 .0
0 .5
1 .0
1 .5
DRatio340/380
la
m
in
e
c
to
m
y
s
h
a
m
s
u
rg
e
ry
0
5 0
1 0 0
1 5 0
t80%(seconds)
6. Figure
10
On
POD7,
capsaicin
evoked
calcium
transients
in
rat
DRG
neurons
as
measured
by
the
ra5o
difference
between
the
peak
ra5o
and
baseline
ra5o
(∆Ra5o
340/380).
Data
are
presented
as
mean±SEM
of
∆Ra5o
340/380.
***p<0.001,
n=40-‐60
cells
from
4-‐6
rats
each
group.
Figure
11
On
POD7,
dura5on
(seconds)
of
recovery
to
80%
of
the
baseline
of
K+
evoked
Ca2+
transients
in
rats
DRG
neurons.
Data
are
presented
as
mean±SEM
of
the
5me
dura5on
to
80%
recovery
of
baseline
of
∆Ra5o
340/380.
***P<0.001,
**p<0.01,
n=
40-‐60
cells
from
4-‐6
rats
each
group.
Figure
12
On
POD7,
percentage
of
capsaicin
responsive
DRG
neurons.
Data
are
presented
as
mean±SEM
of
the
percentage
of
neurons
responsive
to
capsaicin
in
the
whole
DRG.
***p<0.001,
n=
40-‐60
cells
from
4-‐6
rats
each
group.
7. On
pod6
7
there
is
a
sig
diff
between
RTX
and
no
drug
laminectomy
in
running
distance
(two
way
anova,
for
mul5ple
t-‐
test,
day
3,4,5,6,7
is
different.
P=0.016,
0.0013,0.0013,0.0002,0.0004).
On
pod0,
there
is
a
diff
between
RTX
and
no
drug
in
maximum
speed(two
way
anova,mul5ple
t
test,
day
0,1,2,
p=0.0003,0.01,0.02).
there
is
no
diff
between
the
2
group
in
accelera5on.
8. there
is
a
sig
diff
between
RTX
and
no
drug
laminectomy
in
fine
motor
movement
(two
way
anova,
for
mul5ple
t-‐
test,
day
1,2
is
different.
P=0.014,
0.026).
there
is
a
diff
between
RTX
and
no
drug
in
rearing(two
way
anova,mul5ple
t
test,
day
1,
p=0.008).
there
is
no
diff
between
the
2
group
in
accelera5on.
9. Figure
13
On
baseline,
post
RTX
injec5on
day
7,
post
RTX
injec5on
day
14,
thermal
pain
withdrawal
threshold
with
a
cutoff
of
25
seconds.
*p<0.05,
n=4
rats
each
group.
10. Prolonged and enhanced rodent intracellular Ca2+ signal in
dorsal root ganglion after laminectomy was shortened and down
regulated by intrathecal resiniferatoxin
Abstract
Background: postoperative pain is a challenging problem in clinic, compromising
quality of life and leading to extended hospital stay and economic burden and could
develop into chronic pain. Patients report significant pain and hypersensitivity
postoperatively. This study examined the capsaicin and high potassium induced
intracellular calcium response in cultured adult rat DRG neurons on postoperative day
1 and postoperative day 7, and examined the effects of resiniferatoxin treatment on
neuronal intracellular calcium responses and rats behavior including wheel running and
spontaneous home cage activities.
Results: Calcium imaging showed that laminectomy resulted in prolonged duration and
elevated amplitude of high potassium and capsaicin induced DRG neuron intracellular
calcium responses. Also the surgery led to higher responsive ratio in DRG populations,
compared to controls; this was reduced by intrathecal administration of 200ng
resiniferatoxin. Laminectomy resulted in decreased wheel running distance, maximum
speed , home cage rearing and fine movements. Resiniferatoxin partially reversed the
effects of laminectomy on wheel running and home cage spontaneous activities. It also
extended the heat response in hind paw after intrathecal injection.
Conclusions: Laminectomu induces TRP sensitization and higher response to high
potassium in DRG neurons and leads to reduced wheel running and spontaneous
activities in home cage. These effects may be mitigated by preoperative treatment with
11. TRPV1 agonist resiniferatoxin which could lead to desensitization of neurons, increase
wheel running and alleviate spontaneous pain.
Introduction
A large body of evidence point to the importtant role of TrpV1, the capsaicin receptor, in
sensation of noxious heat (Caterina et al., 1997). TrpV1is also believed to play a role in
increased
pain sensitivity following inflammation at least in part because inflammation induced
phosphorylation of TrpV1 results in increased sensitivity of this ion channel to
temperature (Bhave et al., 2002; Cesare et al., 1999; Nilius et al., 2005; Numazaki et
al., 2002). Attention has focused on TrpV1 and TrpV1 cells for their role in pain
sensation and as targets for pain therapies. Notably, TrpV1 antagonists have been
shown to be efficacious in animal models of chronic pain(Szallasi et al.,2007;Wong
andGavva,2009) but, as yet, have not been exploited in human pain management.
An alternate strategy for pain management involves ablation of TrpV1-expressing cells
or TrpV1 fibers. This approach has a long history: more than 25 years ago, capsaicin
injection was reported to dramatically decrease the level of inflammatory mediators
(e.g., substance P) and pain sensation for the life of animals (Jancso et al., 1977; Nagy
et al., 1981; Yaksh et al., 1979). More recently, the potent TrpV1 agonist, resiniferotoxin
(RTX) has been demonstrated to kill TrpV1-expressing neurons by affecting long-term
Ca2+ entry (Olah eal., 2001). RTX has also been used therapeutically in animals and
has been reported to alleviate chronic pain with no effects on discriminative touch or
proprioception (Brown et al., 2005; Karai et al., 2004; Neubert et al., 2003). In a rat
12. model, RTX was shown to eliminate many TrpV1 neurons, dramatically decrease
responses to capsaicin and thermal pain (Karai et al., 2004), and affect
mechanosensation (Tender et al., 2008).
Mishra et al 1
carried out a detailed investigation into the efficacy of RTX in ablating
sensory neurons and demonstrate that although a large subset of TrpV1neurons are
killed by a single injection, other TrpV1 cells are not affected even by repeated RTX
treatments. RTX reduces but does not eliminate capsaicin responses and thermal
sensation of normal mice.
We hypothesized that intrathecal application of RTX may block the DRB neurons
temporarily and reduce the pain behavior. RTX is supposed to work on trpv1 positive
small diameter neurons and leave intact the large diameter axons responsible for
proprioceptive and somatic sensations.The intrathecal approach is proposed to broadly
affect the DRGs surrounding the L4, L5 and their corresponding dermatome including
the laminectomy site to mediate a selective antinociceptive effects. By modifying the
trpv1 positive DRG neurons, this treatment can help relieve different clinical pain
including the postoperative pain.
Here, we set out to determine the function of TrpV1 cells and explored the use of RTX
as a means to affect the intracellular calcium response to capsaicin and hight potassium
stimulations. And to see if it can selectively eliminate trpv1 positive neurons in rats. The
primary aim of this study was to determine whether incisional pain produced a higher
calcium response in DRG neurons and how long it will last. The other aim of the study
was to determine if intrathecal RTX will promote the reduction of calcium response in
these DRG neurons after laminectomy and to check the effect of RTX on the hind paw
13. thermal response and the effect of RTX on wheel running and rodent spontaneous pain
behavior after intrathecal application. Low doses of RTX (100-250ng) has been proved
to be effective when given perineurally2
, in single ganglionic injection or intrathecally3
.
The minimus effective doses should be determined to avoid the po tential side effects
considering the clinical settings. So this study used the 200ng RTX to block multiple
ganglia by applying intrathecally.
Materials and Methods
Animals
Experiments were conducted in adult male Sprague-Dawley rats (Harlan,Indianapolis,
IN, USA) weighing 225-250 g. Animals were housed in groups of 2, maintained in a 12-
hour light-dark cycle with a temperature-controlled environment and given food and
water ad libitum. All procedures used in these experiments were reviewed and
approved by the Institutional Animal Care and Use Committee at the University of Iowa.
Animals were cared for and used in accordance with guidelines of the Committee for
Research and Ethical Issues of the IASP4
.
Chemicals and reagents
1,1'-Dioleyl-3,3,3',3'-Tetramethylindocarbocyanine methanesulfonate (DiI), Fura-2
acetoxymethyl (AM) ester and Pluronic F-127 (stock concentration 20%) were
purchased from Invitrogen (Carlsbad, CA, USA). Concentrations were based on results
from previous studies5
. DiI was dissolved in DMSO (25 mg/ml). Fura-2 AM (2.5µm) was
dissolved in 1mM dimethyl sulfoxide and stored at -20°C. When ready to use, the
Fura-2AM was diluted in bath solution (in mM: 130 NaCl, 5 KCl, 2 CaCl2, 1 MgCl2, 10
14. Hepes, 10 glucose, and pH adjusted to 7.4, and osmolality adjusted with sucrose to
310-325 mOsm), together with Pluronic F-127 (0.025%) immediately before use.
Resiniferatoxin (RTX), (Sigma Aldrich, St Louis, MO, USA) was dissolved in 10/10/80%
mix of ethanol, Tween-80, and saline. Capsaicin (Sigma, St. Louis, MO, USA) was
diluted to a final concentration of 1mM in 1.5% Tween-80, 1.5% alcohol and 97% saline.
Capsaicin (100nM) was diluted in normal bath solution immediately before use. All other
compounds were dissolved in normal bath solution directly. For obtaining high-
potassium extracellular solution (K+
, 30mM), the K+
in normal bath solution was adjusted
to 30 mM by iso-osmotic replacement of Na+
.
Labeling DRG neurons
Prior to tissue harvest (>5 days), sensory neurons innervating the lower back region
were labelled with the retrograde tracer DiI. General anesthesia was induced with
isoflurane (Abbott Laboratories, North Chicago, IL, USA) in room air and maintained at
1.5%, DiI solution (8 µl) was injected onto the right L4 and L5 laminae and surrounding
muscles (4 µl ×2) by positioning the needle 2mm away from the midline (the L4 and L5
spinous processes) with a 30 g injection needle.
Intrathecal injection of RTX or Saline
Under 1.5% isoflurane anesthesia, each rat in the RTX group received intrathecal
(between the L4 and L5 vertebrae) injection of 20 µl RTX (200ng in 20 µl saline) using a
Hamilton syringe and a 25-gauge needle. Each rat in saline group received injection of
20 µl saline.
15. Behavioral pain tests
Rats in the RTX group were tested on baseline, post injection day 7 and post injection
day 14. For withdrawal latency to radiant heat stimuli, unrestrained rats were individually
placed onto a 3-mm thick glass surface (40 × 40 cm) under a plastic chamber (21 ×
27×15 cm) for approximately 15 minutes of acclimation. A focused radiant heat source
was applied from underneath the glass floor onto the center of hind paw. The latency to
evoke withdrawal was determined with a cut-off value of 30 seconds to prevent radiant
heat evoked injury to the hindpaw. Each rat was tested 3 times, with a 5-minute interval
between tests. The mean paw withdrawal latency was determined from 3 tests.
Guarding pain was performed before withdrawal tests to avoid the potential influence of
withdrawal responses on guarding pain.
Wheel running and home cage locomotion tests
Rats were placed individually in cages fitted with exercise wheels (Intellibio Activity
Wheel, Seichamps, France). The rats were housed individually and allow to run on the
exercise wheels with a computer that recorded the distance, speed, acceleration,
revolutions and time (in one hour bins). A stable baseline was achieved after 14 days of
running wheel exposure. The last 3 days were averaged to obtain the baseline. Data
was recorded daily for 14 days. The distance traveled by the rat, acceleration, maximum
speed, total motor movement, and rearing activity of rats became stable after 2 weeks
16. habituation with this experimental protocol. The effects of a treatment on each rat were
analyzed as percentage of the distance traveled during the baseline.
Home Cage Activity Activity during the dark cycle (period when rats are active) after
surgery was monitored. After acclimatization to individual home cages (17 x 5 x 18 inch)
for 3 days, the cages were inserted into a SmartFrame Cage Rack Motor Monitor
System (Kinder scientific, Poway, CA). Two grids of 7x15 infrared beams surrounded
the cage; one was placed 6 cm above the floor to record total locomotor activity as total
beam interruptions. The second series of beams was placed 18 cm above the cage
floor and count rearing, when the rat stood on its hindpaws. Beam breaks were counted
to record total activity and rearing. The data was then transmitted to a host computer
and collected in 1-hour bins. The last 3 days were averaged as the baseline, and then
rats would undergo sham surgery or full laminectomy. We used these monitors to
examine hypermobility by acute pain stimuli.
Laminectomy Procedure
After the induction of general anesthesia with isoflurane in room air and maintained with
1.5% isoflurane. The surgery was performed with spontaneous ventilation through a
nose cone. Each animal was shaved and sterilized with povidone iodine at the incision
site. A 4cm midline longitudinal incision is made with a # 11 blade through the skin
down to the fascia between L3-L6. Blunt dissection was continued through the incision
into the muscle (removing muscle away from bone) to further divide and retracts the
muscle. The posterior bony spine was exposed and L4-L6 spinous processes removed.
The bone between L4-L5 was removed using a battery-powered drill. The dura was
exposed without breaching it. After surgery, the wound was closed in layers; the muscle
17. was closed with 4 sutures, followed by fascia using absorbable 3-0 vicryl suture
(Ethicon, San Angelo, TX, USA). The skin was closed with 4 sutures of 3-0 nylon
(Ethicon, San Angelo, TX, USA).
Cell Culture
Rats were euthanized with 100% carbon dioxide and the right-sided L1-L3 DRG were
surgically removed and desheathed in ice-cold Hanks balanced salt solution (HBSS)
(Gibco, Gaithersburg, MD, USA). DRGs were incubated 20 min at 37°C in 3 ml HBSS
containing 0.125% papain (Worthington, Lakewood, NJ, USA) and then centrifuged at
800 g for 1 min. The papain solution was removed and replaced and incubated for 20
min with 3ml HBSS containing 10 mg collagenase IV (Worthington, Lakewood, NJ,
USA) and 14mg dispase (Invitrogen, Carlsbad, CA, USA). DRGs were then
centrifuged at ∼800 g for 1 min, resuspended with culture media F12 (Gibco,
Gaithersburg, MD, USA). DRGs were mechanically dissociated in this solution with fire-
polished Pasteur pipettes and then plated onto glass coverslips, previously coated by a
solution of 0.1 mg/ml poly-D-lysine (Sigma Aldrich, St Louis, MO,USA) and 5 µg/ml
mouse laminin (BD Bioscience, Bedford, CA, USA). A 0.5-mm-thick silicone ring with a
4-mm-diameter opening sealed to the surface of each well reduced the plating area in
each well. The cells were incubated at 37°C, in 5% CO2 for 2 h. After 2 h, the wells were
filled with 2ml F12 media (Gibco, Gaithersburg, MD, USA) containing 10%(v/v) Fetal
Bovine Serum (FBS), 50 U/ml penicillin , 50 µg/ml streptomycin, and 50ng/ml nerve
growth factor (NGF) (Sigma Aldrich, St Louis, MO,USA), and incubated overnight.
18. Subclassification of DRG neurons
Neurons were divided into small (<30 µm), and large (≥30 µm) based on cell body
diameter as measure with a calibrated eyepiece reticule. Only neurons with visible
nucleus are used. Neurons with a small cell body diameter are likely to be nociceptive
afferents, while those with a large cell body diameter are likely to be non-nociceptive
afferents6
.
Intracellular Ca2+
measurements
All experiments were performed within 24h of tissue harvest. DRG neurons were loaded
with the calcium indicator dye, Fura-2 AM ester (2.5µM) along with 0.025% Pluronic for
20 min at room temperature as previously described 5
. Following fura-2 loading, DRG
neurons were placed in a recording chamber mounted on the stage of an inverted IX-71
microscope (Olympus IX71, Japan) and were continuously perfused for 5 minutes with
normal bath solution before the experiment began. Fluorescence data were acquired on
computer running Slidebook 5.0 software (Molecular Devices, Sunnyvale, CA, USA) at
20°C. Only cells with DiI label were used for calcium imaging. Fluorescence was
alternately excited at 340nm and 380nm using the Lambda DG-4 filter changer (Sutter
Instruments, Novato, CA, USA) via a 10x (Numerical Aperture NA=0.30) or 20x
(NA=0.70) objectives. Emitted fluorescence was collected at 510nm using an ORCA-
03G cooled digital CCD camera (Hamamatsu Photonics, Japan). Pairs of images were
sampled at 1-3 Hz. For each field, an average of 20-40 cells for the 20x objective were
tracked in real time. The ratio of fluorescence emission (510 nm) in response to 340/380
nm excitation was acquired at 1-3 Hz during drug applications. All drugs were applied
19. via a gravity-fed chamber perfusion system. Cells were depolarized by capsaicin (100
nM, 30 seconds) or K+
(30 mM, 30 seconds). Cells not responsive to K+
were excluded.
Two properties of calcium influx were measured: peak ratio (340/380 nm) of the Fura-2
signal, and duration of the Fura-2 signal (calculated as time to 80% of baseline value).
In case of capsaicin depolarization, the percentages of cells responsive to capsaicin
were also compared between groups.
Statistical analysis
Data are expressed as mean± standard error of the mean (SEM) unless otherwise
stated. The “n” for each group refers to the number of neurons. Statistical significance
was determined using Student’s t-test, or ANOVA when appropriate followed by Tukey’s
post-hoc comparisons (GraphPad Prism software, San Diego, CA, USA). Differences at
the P<0.05 level were considered statistically significant.
Results
To assess the effect of laminectomy on the DRG neurons’ intracellular calcium
transients’ responses to K+
or capsaicin induced depolarization, Fura-2 Ca2+
imaging
experiments were performed on DRG neurons that were transiently depolarized with
30nM KCl or 100nM capsaicin. We only analyzed the L1-L3 DRG DiI+ neurons which
were supposed to represent the neurons innervating the laminectomy area at the lower
back region 7
. And only the calcium transients of the potassium responsive neurons
were analyzed.
20. On POD1, laminectomy potentiates K+ evoked Ca2+ transients in DRG neurons
and RTX attenuated the Ca2+ transients enhancement induced by laminectomy.
The magnitudes of K+ evoked Ca2+ transient were analyzed using the change of peak
ratio to baseline. And the decay was measured as time from onset to return to 20% of
peak (T80). Results of this analysis for DiI+ neurons revealed significant effects
associated with laminectomy (P<0.001). When using K+ to depolarize the DRG
neurons, both the magnitude and the decay of Ca2+ transients in DRG neurons were
enhanced by laminectomy. Laminectomy induced higher amplitude (laminectomy
1.70±0.07vs. sham 1.26±0.09, ***p<0.001, Fig.1) and longer decay (laminectomy
269.21±34.98 vs. sham 137.6±15.55, ***p<0.001, Fig.2) when compared with sham
group. Laminectomy significantly enhanced the Ca2+ transients evoked by 30nM KCl
depolarization and attenuated the recovery from depolarization evoked by 30nM KCl.To
determine whether RTX can have effect on DRG neurons’ K+ evoked Ca2+ transients,
we administered 200ng RTX intrathecally at L4 and L5 level 7 days before
laminectomy.Post-hoc (Tukey)analysis indicated that intrathecal 200ng RTX
pretreatment reduced the K+ evoked calcium flux in laminectomy group. (laminectomy
1.70±0.07, vs. RTX and laminectomy 1.14±0.07, *** p<0.001, n= 46 vs 31 cells, 6 rats,
Fig. 1), while the decay of the K+ evoked Ca2+ transient was shortened by RTX
pretreatment (laminectomy 269.2±34.98, vs. RTX and laminectomy 82.43±11.12, ***
p<0.001, n= 25 vs 21cells, 6 rats, ***P<0.001, Fig. 2). Thus, potassium-depolarization
mediated calcium mobilization in laminectomy group was drastically reduced in the
presence of RTX. On POD1, RTX also can reduce the amplitude of calcium flux in sham
rat DRG neurons. (sham 1.26±0.09, vs. RTX and sham 0.88±0.07, **p<0.01, n= 45 vs
21. 44cells, 6 rats, Fig.1). and shortened the duration. (sham 137.7±15.55, vs. RTX and
sham 74.46±8.29, *p<0.05, n= 45,37 cells, 6 rats, Fig.2). Therefore, on POD1, in both
laminectomy rats and sham rats, RTX reduced the calcium response to KCl , either by
reducing the amplitude of calcium peak or by shortening the decay of the transient.
On POD1, capsaicin evoked Ca2+ transients in DRG neurons was also potentiated
by laminectomy, which was attenuated by intrathecal RTX.
Laminectomy induced higher amplitude when compared with sham group (laminectomy
2.60±0.14 vs. sham 2.03±0.14, *p<0.05, Fig. 3 Post hoc analysis (Tukey test)) and slow
the recovery of transients in small diameter neurons (laminectomy 986.10±116.10, vs.
sham 511.31±78.65, ***P<0.001, Fig. 4). And there is also a significant increase in the
percentage of capsaicin responsive neurons in the DRG populations induced by
laminectomy (laminectomy 75.20±3.79% vs. sham 63.67±1.93%, *P<0.05, Fig. 5).
As observed RTX effect on potassium induced calcium transients, there was also a
statistically significant effect of pre-surgery RTX in both the magnitude (laminectomy
2.60±0.14, vs. RTX and laminectomy 1.52±0.16, *** p<0.001, n= 40 vs 34 cells, 6 rats,
Fig. 3)and decay (laminectomy 986.10±116.10, vs. RTX and laminectomy
378.70±69.18, *** p<0.001, n= 32 vs 27 cells, 6 rats, Fig. 4) of the capsaicin evoked
Ca2+ transient in small DiI+ DRG neurons. In addition, RTX decreased the percentage
of capsaicin responsive DRG (laminectomy 75.20±3.79% vs. RTX 51.00±3.67%,
***p<0.001, Fig. 5). It also decreased the percentage of capsaicin responsive DRG in
sham rats (sham 63.67±1.93% vs. RTX and sham 33.10±1.78%, ****P<0.0001).
22. On POD1, in both laminectomy rats and sham rats, RTX reduced the calcium response
to capsaicin, either by completely eliminating the transient or by reducing the amplitude
of calcium peak or by shortening the decay of the transient.
On POD7, laminectomy did not produce significant changes in K+ evoked Ca2+
transients in DRG neurons either in amplitude (laminectomy 1.04±0.11vs. sham
1.18±0.08, p>0.05,n= 23 vs 40 cells, 4 rats Fig. 6) or decay(laminectomy 108.70±26.31
vs. sham 116.2±12.79, p>0.05, Fig. 7).
On POD7, laminectomy extended the decay of capsaicin evoked Ca2+ transients.
And RTX attenuated the laminectomy enhanced capsaicin response in DRG
neurons
On POD7, even the magnitude of capsaicin evoked Ca2+ transients did not differ
between laminectomy group and sham group(laminectomy 2.19±0.18 vs. sham
1.77±0.15,p>0.05, n=20 vs 38 cells, 4 rats each group Fig. 8) although RTX decrease
the amplitude both in laminectomy rats (laminectomy 2.19±0.18, vs. RTX and
laminectomy 1.21±0.12, n=20 vs 37 cells , 4 rats each group,*** p<0.001, Fig. 8) and
sham rats(sham 1.77±0.15% vs RTX and sham 0.85±0.09%, n=38 vs 30 cells, 4 rats
each group, ***P<0.001, Fig. 8).
However there was still increased decay of capsaicin evoked Ca2+ transients in
laminectomy group (laminectomy 1044±149 vs sham 414±44.76 seconds, n= 25vs 33
cells, 4 rats each group, ***P<0.001, Fig. 9), and RTX shortened the decay in both
laminectomy rat (laminectomy 1044.0±149.3 vs RTX and laminectomy 188.6±33.68
23. seconds, n=25 vs 26 cells, 4 rats each group, ****P<0.0001, Fig. 9) and sham
rats(sham 414±44.76% vs RTX and sham 161.2±11.89%, n=33 vs 35 cells, 4 rats each
group, *P<0.05, Fig. 9).
On POD7, there was no significant difference in terms of the percentage of capsaicin
responsive neurons between the sham group and laminectomy group (laminectomy
71.75±6.96% vs. sham 59.51±2.82%, 4 rats each group, p>0.05, Fig. 10), even when
RTX still lowered the percentage of capsaicin responsive DRG neurons both in RTX
pretreated laminectomy group (laminectomy 71.75±6.69% vs RTX laminectomy
45.0±2.04%, 4 rats each group,**p<0.01, Fig. 10) and sham rats(sham 59.51±2.82% vs
RTX and sham 33.60±1.50%, n=10 vs 5 rats each group, *P<0.001, Fig. 10).
Thus, on POD7, RTX still attenuated the laminectomy enhanced DRG neurons’
responses to capsaicin.
Effects of RTX on wheel running and home cage activity
We observed the whole week after laminectomy using RTX. And found significant
changes after RTX administration. The attenuated behavior was recovered faster
compared with laminectomy alone. Including running distance and locomotion.
24. rtx la m i D is ta n c e
B
a
s
e
lin
e
0
1
2
3
4
5
6
7
0
5 0
1 0 0
1 5 0
2 0 0
R T x + L a m in e cto m y lia n g
N o D ru g (N o A lz e t) L a m i lia n g
S h a m lia n g
rtx la m i M a x S p e e d
B
a
s
e
lin
e
0
1
2
3
4
5
6
7
0
5 0
1 0 0
1 5 0
R T x + L a m in e cto m y
L a m i
S h am
wo
On
pod6
7
there
is
a
sig
diff
between
RTX
and
no
drug
laminectomy
in
running
distance
(two
way
anova,
for
multiple
t-‐
test,
day
3,4,5,6,7
is
different.
P=0.016,
0.0013,0.0013,0.0002,0.0004).
On
pod0,
there
is
a
diff
between
RTX
and
no
drug
in
maximum
speed(two
way
anova,multiple
t
test,
day
0,1,2,
p=0.0003,0.01,0.02).
there
is
no
diff
between
the
2
group
in
acceleration.
25. R T X F in e m o to r m o v e m e n t
B
a
s
e
lin
e
0
1
2
3
4
5
6
7
0
5 0
1 0 0
1 5 0
S h am
L a m in e c to m y
L a m in e cto m y -R T x
R T X R e a rin g
B
a
s
e
lin
e
0
1
2
3
4
5
6
7
0
5 0
1 0 0
1 5 0
S h a m lia n g
L a m in e c to m y
L a m in e cto m y R T x
26. there is a sig diff between RTX and no drug laminectomy in fine motor movement (two
way anova, for multiple t- test, day 1,2 is different. P=0.014, 0.026). there is a diff
between RTX and no drug in rearing(two way anova,multiple t test, day 1, p=0.008).
there is no diff between the 2 group in acceleration.
Effects of RTX on the thermal hyperalgesia
The effects of the intrathecal administration of RTX (200ng) on thermal hyperalgesia in
naïve rats at 7 days and 10 days after injection of RTX were investigated.
Our results show that the intrathecal administration of 200ng RTX inhibited the thermal
response of hindpaw (Fig. 11), which confirmed the valid intrathecal injection of RTX.
Discussion
The pain after operation represents a significant clinical challenge for treatment. The
search for new treatment options of postoperative pain has been directed toward
specific mediators and modulators of pain and inflammation.
In this study, we showed that laminectomy sensitized the DRG sensory neurons
indicated by elevated amplitude of calcium response curve and prolonged duration both
in response to high K and capsaicin and a higher percentage of capsaicin responsive
,TRPV1 positive neurons.
The present study also showed that it RTX attenuated DRG responsiveness to
potassium and capsaicin evidenced by recovered amplitude, reversion of prolonged
duration to high K and capsaicin and reduced percentage of TRPv1 positive DRG
neurons.
27. Furthermore, on pod7 laminectomy still has prolonged the duration of intracellular
neuronal calcium response to capsaicin, which is also reversed by pretreatment of i.t.
RTX indicated by a normalized duration of calcium response. This showed an extended
effect of RTX.
Laminectomy produces a decrease in rat wheel running and spontaneous home cage
activities. I .T. RTX pretreatment reversed the decrease of wheel running distance and
recovered the rearing and fine movement. Because there were no sided effects
observed in the experiment, thus the safety profile of RTX was established. We propose
that RTX intrathecal application could be a useful strategy in alleviating postoperative
pain.
We developed an rat model of postoperative pain and examined whether TRPV1 play a
role in mediating postoperative pain injury. In sensory dorsal root ganglia, the trpv1 are
expressed on small and medium –sized neurons and are mainly involved with mediation
of nociceptive information. TRPV1 is expressed by the type II A-delta and C-fiber
neurons, functioning as a molecular integrator of nociception. TRPV1-positive neurons
are essential for the development of mechanical allodynia.
Trvp1 is sensitized by phosphorylation of trpv1 via cAMP8
. and we tested he effect of
specific TRPV1 agonists could overexcited DRG neurons and provide a potential way of
relieving pain.
Prior studies have shown that pain could induce expression of TRPV1 and neuron
phenotype change. Nerve injury and inflammation could enhance the expression of
28. trpv1 including the transcription, translation, post translational modification and
trafficking of trpv1(Hucho T 2007 neuron, zhang x, EMBO J 2005, Ji RR, Neuron 2002).
The net effect is central and peripheral sensitization. This could switch the phenotype of
DRG nruosns and spinal cord neurons from naïve state to an altered state. Thus
redcuce the pain threshold(Patapoutian A, 2009).Trpv1 could enhanced form gene
level, translational level (mRNA), and post-translational modifications such as
phosphorylation, n-glycosylation and covalent modification9
. Especially phosphorylation
by PKCᵋ (Cesare 1999, Mandadi S 2006 pain) 10
and PKA(Bhave 2002, Lopshire, 1998).
Also there is evidence showing the phosphorylation of Trpv1 is enhanced. .Inflammaorty
mediators sensitized trpv1 through phosphorylation of trpv1 by protein kinase PKCᵋ
(Mandadi S 2006 Pain zhang H, 2007 J neuroscience).
In postoperative rat, trpv1 could be elevated because of gene level, rna level and
posttranstlational level change. Recent study demonstrated that TRpv1 antagonist
intratheally and local application both can allevievate thermal hyperalgsiea and
mechanical allodynia(Uchytilova mole pain ,2014 pure behavior). Also in a model of
thoracotomy postoperative pain, RTX significantly reduced mechanical allodynia
following surgery.(Shin Jw , 2010). TRPV1-expressing C-fibers are sensitized to heat
and acid after incision (Kang S 2010 Pain), All this indicated the involvement of trpv 1 in
postoperative pain. As we have demonstrated here, there were marked differences in
small DRG neurons between laminectomy group and sham group with respect to the
magnitude and decay of capsaicin-evoked Calcium transients. In order to identify
neurons innervating the site of surgery, DiI was injected in lumbar region prior to the
29. surgery. One day after surgery, there was a significant increase in the magnitude and
decrease in the decay of capsaicin evoked calcium transients in small DRG neurons.
Similar decreases in decay were also observed in the transients evoked by capsaicin
even 7 days after laminectomy. The reason of the enhanced intracellular calcium
transients during inflammation could be the inflammatory mediators such as nerve
growth factor because they might affect the calcium efflux, uptake and binding of
calcium and thus contribute to the enhanced calcium transients during inflammation (Lu,
2008 ). The trpv1 sensitaztion in laminectomy group could be attributed to inflammatory
mediators such as Nerve growth factor (NGF) because it induces sensitization of the
TRPV1(Zhu W, 2007 mol cell neurosci). Other Inflammatory mediators (BK, substance
P) also modulate TRPV1 activity (Sugiura T. Bradykinin lowers the threshold
temperature for heat activation of vanilloid receptor
1. J Neurophysiol 2002; 88:544–8. Neurokinin-1 receptor enhances TRPV1 activity in
primary sensory neurons via PKCepsilon: a novel pathway for heat hyperalgesia. J
Neurosci. 2007 Zhang H). Studies have shown that trpv1 sensitization(by heat)
produced a larger membrane deploarizations in rat DRGs. This could lower the
threshold and increase excitability of DRG neurons. (Neelands, 2008 inflam research,
zhang xf, 2011). We demonstrated that surgery induced an enhanced calcium response
to both potassium and capsaicin. Because Intracellular calcium plays a critical role in
mediation of many cellular processes such as transcription, ion channel activity, it
regulates the neuronal excitability and transmitter release (Berridge, 2005)and hormone
release, and Ca2 +
-dependent gene transcription. , so the enhanced calcium response
seen in small DRG after surgery might contribute to the postoperative pain by changing
30. afferent excitability and facilitating transmitter transcription and release from the
nociceptive afferents. Indeed, pervious study showed that activation of trpv1 in afferent
terminals could increase the rate of glutamate release by allowing calcium into neurons
(Shoudai 2010). Such results suggest that activated, presynaptic TRPV1+ receptors
trigger continuous resting release of glutamate vesicles at physiological temperatures
only in capsaicin-responsive terminals. Thus the sensitization of DRGs found in
postoperative pain model indicated the higher excitability of DRG neurons and
increased neurotransmitter release in afferent terminals.
In terms of percentages of trpv1 response positive cells, unregulated trpv1
responsiveness in primary-afferent fibers occurs in inflammatory disease and irritable
bowel syndrome (yiangou 2001, Chan 2003). Perivous study has shown that nerve
injure leads to immnunoreactivity changes from trpv1 negative to positive in large
diameter neurons after nerve injury(Hudson 2001). A similar gain of capsaicin sensitivity
was seen in calcium imaging recently after nerve injury (Kim 2014 neuron). These
phenotypic switch was mostly seen in nerve injury and chronic pain(Costigan M 2009
Annu Rev Neurosci). Interestingly, cancer pain also leads to increase of trpv1
expression in both myelinated DRG neurons and perptidergic unmyelinated DRG
neurons (Niiyama Y, 2007 neuroscience). In our experiment, the intracellular calcium
response to capsaicin is also enhanced in terms of percentages of positive cells. The
percentage of TRPV1 responsive DRG rose in pain group was higher compared with
control group. The percentage change might reflect a phenotype change in DRG
population. This findings extended previous research results by showing that
31. unregulated trpv1 responsiveness in primary-afferent fibers occurs in postoperative
pain,
Together, these data are consistent with the hypothesis that the TRPV1 contribute to
postoperative pain DRG excitability. Indeed, the responses are both elevated in pod1
and pod7 when stimulated with capsaicin. To further understand the role to trpv1-
positive neurons in development of postoperative pain, we studied the Pod7 response in
DRGS. On PoD7, We found the the amplitude is reduced and d the percentage is
recovered. The amplitudes of capsaicin evoked calcium transients and percentages of
capsaicin responsive neurons are not higher in surgery group when compared with
control. This showed that the surgery induced a recoverable enhanced response in
DRGS indicating the disappearance of the inflammaotory part of postoperative pain.
However on pod7 the duration of capsaicin induced calcium response remains longer
in surgery group. This could due to the long- lasting remaining sensitization of DRGS
after surgery. This extended effect of surgery on calcium responses was reflected by
our wheel running results, which showed that on Pod7 there is still a difference between
RTX and no treatment group in running distance(On pod6 and pod7 there is a sig diff
between RTX and no drug laminectomy in running distance (two way anova, for multiple
t- test, day 3,4,5,6,7 is different. P=0.016, 0.0013,0.0013,0.0002,0.0004).
In our study, One day after the laminectomy, there was also a significant increase in the
magnitude and decrease in the decay of potassium evoked calcium transients in DiI
labeled neurons. We found the response duration, amplitude of DRG response to k30
to be elevated on POD1. And this enhanced response disappeared on POD7. Evidence
32. showed that the nerve injury in DRG leads to decrease in resting calcium concentration
(Fuchs ,2005) and a decrease in both the magnitude and decay of evoked calcium
transients (Fuchs 2007)and nerve injury also leads to a decreased voltage gated
calcium currents in injured DRG (MaCallum 2003, Baccei and Kocsis,2000). Also there
was reports that inflammation is associated with increases in magnitude and decrease
in decay of the high potassium evoked transient in small putative nociceptive
afferents(Lu 2008,neuroscience). In the present postoperative pain model, high
potassium induced weak excitation of naïve DRG , but strongly activate the DRG in
laminectomy group as indicated by increase in calcium transients’ amplitudes and
durations. This augmented responsiveness indicated neuronal sensitization. This
stronger activation is possibly due to trpv1 sensitization in laminectomy group because
inflammatory mediators induces an acute sensitization of nociceptive DRG neurons
partially through sensitization of the TRPV1(Zhu W, 2007 mol cell neurosci). The high
responsiveness to KCl and capsaicin shown in our postoperative pain model support
the idea of sensitization of DRG and phenotypic changes in DRGs after surgery. This
enhanced responsiveness to potassium disappeared on POD7.
Using this pain model, we also showed that the significantly augmented DRG neurons’
response to both potassium and capsaicin challenges are blunted by the application of
TPV1 agonist RTX. Also the percentage changes happened following the surgery are
also prevented by pretreatment of RTX. RTX mediated antinociceptive effects both in
home cage locomotion and wheel running. Trpv1 agonists-induced analgesia is
proposed be due to receptor desensitization because of a sustained activation of
33. receptors()Jeffry Jap los one 2009, Raisinghani M 2005 j physiol). I. t. RTX couls cause
aa sustained increased freuqencey of spontaneous EPSCs and lead to a
desensitization response and depolarization block, synaptic failures(Jeffry Ja plosone
2009 sikand p, 2007 j physiol).. RTX has been shown to cause both spinal cord and
DRG changes if applied intrathecal.
RTX could induce a degenaeraion of trpv1 positive neurons by causing a neurotoxic
influx of calcium ion through the channels resulting a calcium induced neuronal
death(karai 2004). Our data extended the previous findings by showing that the
percentage of DRG neurons responveis to capsicien is significantly reduced aftet RTX
treatment. Also it has been shown that the ablation of central terminals of trv1
expressing neurons occured. after i.t. RTX (Bishinoi M 2011 j pain). Even when used
perineurally and remote from the neuronal perikary in DRG , RTX still could ablate DRG
TRPV1 positive neurons (Neubert 2008). RTX is neuroexcitotoxic and ablate neurons.
RTX is a promising drug for treatment because of its desensitizing effect and ablate
effect. Single dose might desensitize ,repeated dose or high dose could cause
neurotoxity and permanently degenerate nerve Trpvv1 + nerve endings (Ladarola
2011). RTX induced a prolonged increase in intracellular calcium in Trvp1 positive
neurons in human dorsal root ganglion cultures, , while leaving other adjacent neurons
unaffected(Karai, 2004). It has been proved that intrathecal RTX could reduce TRpv1,
CGRP and trpa1 immunoreactivities in the lumbar dorsal root ganglia and (karai 2004,
Neubert 2008 Klafke 2012 neuroscience), indicating the deltetion ot DRG TRPV1-
expresssing neurons(Neubert). Intrathecal rtx also produce ablation of central nerve
34. terminals of DRG neurons indicated by decreased immune reaction of TRV1 in spinal
cord dorsal horn (Bishnoi 2011). This cellular changes were reflectd in behavioral tests
such as prolonged hind paw thermal stimulation latency and carrageenan-induced
inflammatory therma lanalgesia(Bishnoi 2011). Our thermal test on hind paw confirmed
the prolonged radiant noxiouls thermal stimulus latency and suggested the specific
deletion of trpv1 positive neurons. In the present study, intrathecal RTX reduced the
calcium response. We found that RTX producing a significantly reduced responsiveness
to potassium and capsacin in DRGs.Also In calcium imaging, the percentages of trpv1
responsive DRG neurons decrease after RTX application both in postoperative rats and
sham rats further proved that the intrathecal RTX targeted trpv1 positive neurons in
DRG and made them inert to high potassium and capsaicin. We propose that i.t. RTX
also affect DRG trpv1 channels because RTX diffused to DRG and act on primary
afferent neurons. This may partly explained our finding that after rtx, the DRG response
to capsaicin are reduced.
This further extended the findings that RTX decreasing the TRPV1 immunoreactivities
in DRG after both perineural and intrathecal RTX application(neubert Jk, 2008, Klafke
2012 neuroscience)
RTX has been used for debilitating arthritic and cancer pain . In rats of postoperative
pain, RTX can significantly reduce the pain(my paper). Also the RTX has been shown to
be effective in treatment of neuropathic pain(gariel Tender, spinaljournal 2008) because
RTX jinjection acuse an excitotoxic effect when injected into sensory ganglia. RTX is a
35. transient receptor potential vanilloid 1 channel agonist. It also contributes to
inflammatory and heat allodynia relieve. Our intrathecal injection of RTX also cause an
inert DRG neuron reaction to K30 and capsaicin, both on pod1 and pod7. Extensive
research dta have shown that trpV1 positive neurons mediate thermal hyperalgesia in
pain models.(11-16 of Tender 2008 spine ournal). Moreover , the same cells apppeart
to be responsible ofre the mediation of neurogenic inflamesion(13.17-19), blockage of
trpv1 apears eefective in reversing established hyperalgesia associate with
inflammation(18). Our data corroborated that RTX could abolish the heat induce pain in
rats, evidence by the extenede time response time to the heat application on hindpaw in
rats treated with i.t. RTX. The paw became less responsive to heat. These results are in
concordance with previous studies showing theat trpv1-posticitve neurons mediated
termal hyperalgesia.
Also it is shown that inhibition of trpv1 centrally could inhibit the mechanical
hyperalgesia even it is not a mechanotransducer(Kim 2014 Neuron). So far there have
been no data to demonstrate the involvement of trpv1-positive neurons in mechanical
sensitivity or allodynia. However, some earlier reports showed that RTX injected in
DRGs could elevated the average withdrawal threshold of the paws and the magnitude
of touch –evoked allodynia was improved, which means the RTX improved tactile
allodynia via selectively eliminating the trv1-positive cells in the DRGs. This could be
explained by the cross-talk between A-beta, A-delta and C-fiber neurons in DRGs (23 of
36. tender 2008) . After operation, large amounts of glutamate and aspartate are released
into extracellular space and the inhibitory neurotransmitter GABA is reduced in spinal
dorsal horn (22) and also in the DRGs, resulting the hypersensitivity of A fibers and C
fibers. When use a touch test of allodynia, the A-beta fibers carried the signals to the
DRG and excited not only the A-beta cell bodies but also A-delta and C neurons
because of the cross-excitation. The excitation of a A-delta and C neurons subsequently
contribute to the touch induced pain and personal’s experience and emotional status.
Thus, deletion of trpv1 neurons (include A-delta and C fibers) could reduce the
excitation of A-delta and C- fibers in DRGs and thus reduce the whole animal’s
response to touch.
Also even we did not test the mechanical threshold of back region. There was evidence
showing that the surgery affected the cutaneous sensitivity in the back region after
thoracotomy and RTX could relieve the allodynia developed after(Shin 2010) surgery11
.
It can relieve pain like shown in behavioral tests wheel running and spontaneous
activities. It already has been shown when used in large doses intrathecally(3.8ug/kg), it
is able to alleviate capsaicin induced nocifensive behavior(bishnoi 2011 j pain) and both
capsaicin and carrageenan induced inflame thermal hypersensitivity
(bishnoi 2011 j
pain).
Also it is shown that inhibition of trpv1 centrally could inhibit the mechanical
hyperalgesia even it is not a mechanotransducer(Kim 2014 Neuron).
We observed the spontaneous activity after surgery and the wheel running behavior
after laminectomy instead, it is shown that the wheel running and home cage activity are
changed by laminectomy and recovered partially due to RTX. We say the RTX
37. improved the running distance and the spontaneous activity in postoperative rats. Such
as rearing and fine movements in homecage, which proved that RTX mediated
antinociceptive effects and even after intrathecal RTX, animal maintain the motor
control. There was report that RTX caused a long lasting analgesia in cancer pain
model even up to 14 week after application and the hind paw heat response was
significantly reduced (Brown 2005). Our heat test further confirmed that intrathecal RTX
induced a higher heat response threshold. Also it has been shown that lumbar epidural
application of RTX resulted a focused distribution to lumbar neurons and lead to a
profound and segmental analgesia12
. Recently it was shown that intrathecal RTX could
lead to dose-dependent deletion of TRPV1 CGRP and phosphor ERK
immunoreactivities in superficial spinal cord such as laminae I and II. Other studies
showed the physiological sensation could recover 2 weeks after RTx treatment2
.
In summary, the high responsiveness to capsaicin and potassium in DRG reflected the
sensitization of neurons and this indicates high excitability and more neurotransmitter
release in peripheral nociceptive neurons. RTX is a promising pain killer because both
the calicium imaging and behavioral test support its use in postoperative pain.
1.
Mishra
SK,
Hoon
MA:
Ablation
of
TrpV1
neurons
reveals
their
selective
role
in
thermal
pain
sensation.
Mol
Cell
Neurosci
2010;
43:
157-‐63
2.
Neubert
JK,
Mannes
AJ,
Karai
LJ,
Jenkins
AC,
Zawatski
L,
Abu-‐Asab
M,
Iadarola
MJ:
Perineural
resiniferatoxin
selectively
inhibits
inflammatory
hyperalgesia.
Mol
Pain
2008;
4:
3
38. 3.
Karai
L,
Brown
DC,
Mannes
AJ,
Connelly
ST,
Brown
J,
Gandal
M,
Wellisch
OM,
Neubert
JK,
Olah
Z,
Iadarola
MJ:
Deletion
of
vanilloid
receptor
1-‐expressing
primary
afferent
neurons
for
pain
control.
J
Clin
Invest
2004;
113:
1344-‐52
4.
Zimmermann
M:
Ethical
guidelines
for
investigations
of
experimental
pain
in
conscious
animals.
Pain
1983;
16:
109-‐10
5.
Lu
SG,
Zhang
X,
Gold
MS:
Intracellular
calcium
regulation
among
subpopulations
of
rat
dorsal
root
ganglion
neurons.
J
Physiol
2006;
577:
169-‐90
6.
Waxman
SG,
Dib-‐Hajj
S,
Cummins
TR,
Black
JA:
Sodium
channels
and
pain.
Proc
Natl
Acad
Sci
U
S
A
1999;
96:
7635-‐9
7.
Takahashi
Y,
Chiba
T,
Kurokawa
M,
Aoki
Y:
Dermatomes
and
the
central
organization
of
dermatomes
and
body
surface
regions
in
the
spinal
cord
dorsal
horn
in
rats.
J
Comp
Neurol
2003;
462:
29-‐41
8.
Bhave
G,
Zhu
W,
Wang
H,
Brasier
DJ,
Oxford
GS,
Gereau
RWt:
cAMP-‐dependent
protein
kinase
regulates
desensitization
of
the
capsaicin
receptor
(VR1)
by
direct
phosphorylation.
Neuron
2002;
35:
721-‐31
9.
Voolstra
O,
Huber
A:
Post-‐Translational
Modifications
of
TRP
Channels.
Cells
2014;
3:
258-‐87
10.
Cesare
P,
Dekker
LV,
Sardini
A,
Parker
PJ,
McNaughton
PA:
Specific
involvement
of
PKC-‐epsilon
in
sensitization
of
the
neuronal
response
to
painful
heat.
Neuron
1999;
23:
617-‐24
11.
Shin
JW,
Pancaro
C,
Wang
CF,
Gerner
P:
The
effects
of
resiniferatoxin
in
an
experimental
rat
thoracotomy
model.
Anesth
Analg
2010;
110:
228-‐32
12.
Szabo
T,
Olah
Z,
Iadarola
MJ,
Blumberg
PM:
Epidural
resiniferatoxin
induced
prolonged
regional
analgesia
to
pain.
Brain
Res
1999;
840:
92-‐8