1. Introduction
Methods
Abstract
Skeletal muscle possesses the ability to repair and regenerate following injury. The
inflammatory response is an essential component of muscle repair. At critical periods
following injury, macrophages switch from a pro-inflammatory (M1) to a regenerative
(M2) phenotype to aid in the repair. We hypothesize that Klotho, a protein expressed
by inflammatory cells within injured muscle, is an important mediator of macrophage
phenotype. Wild-type (WT) mice were compared to a Klotho deficient (KL.Hm)
strain. Following a sterile BaCl2 injury, muscles were analyzed at 7 and 21 days post
injury (PI). Antibodies to CD68 (M1 marker), CD163 (M2c marker), CD206 (M2a
marker) were used to evaluate macrophage phenotype. Following injury, ablation of
Klotho increased CD68+ cells 7 days PI and decreased CD163+ and CD206+ cells 21
days PI compared to WT. In addition to macrophage phenotype, we assayed for the
presence of eosinophils using anti-MBP-1 and regulatory T-cells using anti-FoxP3
because they are also leukocytes that can affect muscle regeneration. At 7 days PI,
MBP-1+ cells were down regulated in KL.Hm compared to WT. There were no
differences in FoxP3+ cells after injury. Our results reveal that Klotho deficiency
promotes a pro-inflammatory environment after muscle injury. Therefore, Klotho may
be an important mediator of muscle regeneration by affecting the inflammatory
response following injury.
Martina M. Shoukralla, Steven S. Welc, and James G. Tidball
Department of Integrative Biology and Physiology, University of California Los Angeles
Discussion and Conclusions
Klotho’s Effects on Skeletal Muscle’s Inflammatory Response During Muscle Regeneration
Results
• Mouse Strains: heterozygous Klotho mice on C57BL/6 background were provided by Dr. M.
Kuro-o at the University of Texas Southwestern Medical Center as a gift.
• BaCl2 sterile muscle injury: The left tibialis anterior (TA) was injected with 50 µl of a 1.2%
barium chloride (BaCl2) solution. The contralateral leg served as an uninjured control muscle.
Mice were sacrificed at 7 and 21 days post injury.
• Immunohistochemistry: Rat αCD68 (1:100, Serotec) was used as a primary antibody for
labeling M1 macrophages and rabbit αCD163 (1:100, Santa Cruz), and rat αCD206 (1:50,
Serotec) were used as primary antibodies for identifying M2 macrophages. Anti-major basic
protein (αMBP-1) (1:3000) was used as a primary antibody to label eosinophils and αFoxP3
was used to label regulatory T cells.
1. Inflammation is an important step in skeletal muscle regeneration.
2. Following injury pro-inflammatory Th1 response takes place. This period is marked
by the infiltration of neutrophils and M1 macrophages.
3. The inflammatory phase resolves into a pro-regenerative Th2 phase several days
after injury. A switch in macrophage phenotype from M1 to M2 highlights this phase.
• Klotho is an important protein for regulating the inflammatory environment after muscle
injury.
• Klotho-deficient mice experienced an increase in M1 macrophages and a decrease in
eosinophils at 7 days PI.
• The reduction in M2 macrophages in Klotho-deficient mice at 21 days PI suggests
impairment of the Th2 inflammatory response that is associated with muscle
regeneration.
• Additional work is needed to determine if Klotho is a direct or indirect regulator of
macrophage phenotype.
• Overall, our data support the possibility that Klotho has a significant role in regulation
muscle inflammation and repair following injury.
• Tidball JG. Mechanisms of muscle injury, repair, and regeneration. Compr Physiol. 2011 Oct;
1(4):2029-62. doi: 10.1002/cphy.c100092. Review.
Figure 2: Regulatory T-cell abundance in KL.Hm vs. WT.
Non-injured, 7 days PI and 21 days PI TA muscles were analyzed for regulatory T-cell (T-
reg.) abundance. FoxP3 is a T-reg. marker. Data are presented as mean ± SEM. One-way
analysis of variance was used to test whether differences between groups were significant at
p < 0.05. Significant differences between groups were identified using Bonferroni's follow
up test. # vs. non-inj. of same genotype.
A.
B.
C.
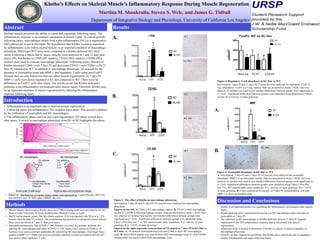
Figure 1: The effect of Klotho on macrophage phenotype.
Non-injured, 7 days PI and 21 days PI TA muscles were analyzed for macrophage
phenotype.
Figures on the left: A. CD68 is M1 macrophage marker. B. CD163 is M2c macrophage
marker. C. CD206 is M2a macrophage marker. Data are presented as mean ± SEM. One-
way analysis of variance was used to test whether differences between groups were
significant at p < 0.05. Significant differences between groups were identified using
Tukey’s Post Hoc test. * vs. WT injured under same conditions. # vs. non-inj. of same
genotype. $ vs. 7-d P.I. of same genotype.
Figures on the right represent cross-sections of TA muscle at 7 days PI in KL.Hm vs.
WT mice. A. TA muscles immunolabeled with anti-CD68 to show M1 macrophages
(red). B. Anti-CD163 marker was used to show M2c macrophages (red). C. Anti-CD206
immunolabel was used to show M2a macrophages (red).
CD68
Non-inj 7d PI 21d PI
0
10000
20000
30000
cells/mm3
WT
KL.Hm
#
#
*
$
$
CD163
Non-inj 7d PI 21d PI
0
2000
4000
6000
cells/mm3
WT
KL.Hm
#
CD 206
Non-inj 7d PI 21d PI
0
5000
10000
15000
20000
cells/mm3
WT
KL.Hm
#
MBP
Non-inj 7d PI 21d PI
0
1000
2000
3000
4000
5000
cells/mm3
WT
KL.Hm
#
*
#
$
Figure 3: Eosinophil abundance in KL.Hm vs. WT.
A. Non-injured, 7 days PI and 21 days PI TA muscles were analyzed for eosinophil
abundance. MBP-1 is an eosinophil marker. Data are presented as mean ± SEM. One-way
analysis of variance was used to test whether differences between groups were significant at
p < 0.05. Significant differences between groups were identified using Tukey’s Post Hoc
test. * vs. WT injured under same conditions. # vs. non-inj. of same genotype. $ vs. 7-d P.I.
of same genotype. B. Cross-sections of TA muscle, at 7 days PI, immunolabled with anti-
MBP-1 to show eosinophils (red).
0
200
400
600
cells/m3
FoxP3, WT vs KL.Hm
Non-inj 7d PI 21d PI
#
WT KL.Hm
A.
B.
WT
KL.Hm
KL.Hm
WT
WT
KL.Hm