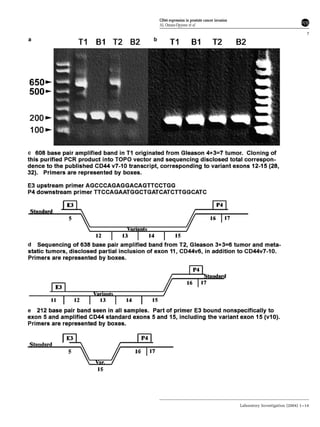
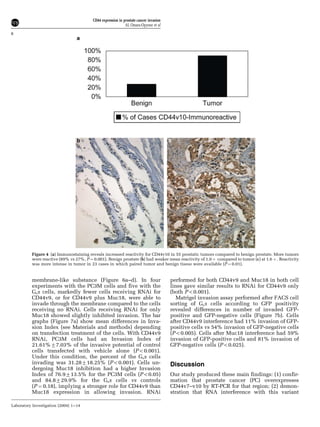
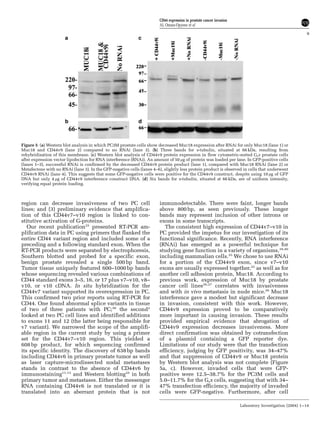
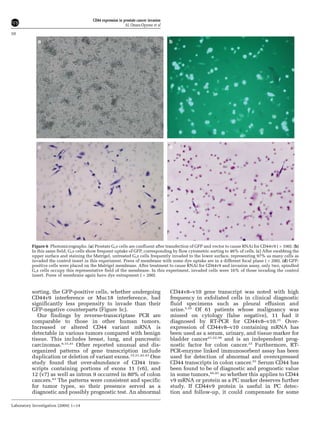
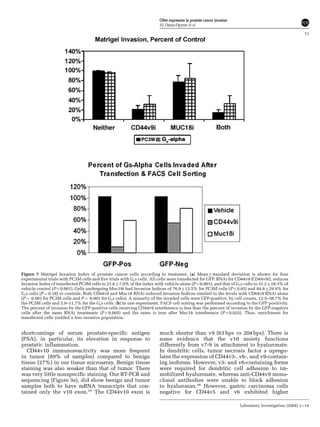
This document summarizes a study examining the role of CD44 variant 9 (CD44v9) and Muc18 expression in prostate cancer invasion and metastasis. The study found that prostate cancer tissues and metastases overexpressed CD44v7-v10 isoforms compared to benign tissue. Silencing CD44v9 expression in cultured prostate cancer cells significantly reduced invasiveness into Matrigel, while silencing Muc18 had a smaller effect. A more invasive cell line, Gsa, was also found to overexpress CD44v9. The results suggest that prostate cancer invasion is more influenced by CD44v9 expression than by Muc18 expression.