Syllabus
• Introduction
• Definitionand Scope of Microbial
Physiology
• Phylogenetic Tree of Life
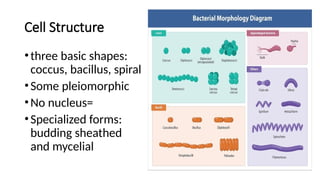
• Bacterial Cell
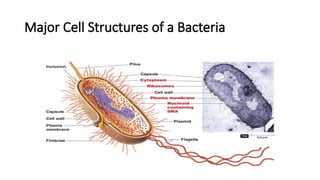
• Major Cellular Structures
• Chemistry and Synthesis of Cellular
Structures
• Microbial Growth
• Definition of Growth
• Measurement of Growth
• Growth Physiology
• Factors Affecting Growth and Stress
Responses
• Microbial Nutrition and Solute Transport
• Nutritional Reqs of Mcgs
• Mechanisms of Transport
• Bioenergetics in the Cytosol
• High Energy Molecules
• Mechanisms of ATP Synthesis
• Prokaryotic Metabolism
• Central Metabolic Pathways
• Fermentation
• Photosynthesis
• Metabolism of Lipids, Nucleotides, AA, and
Hydrocarbons
• Inorganic Metabolism
• C1 Metabolism
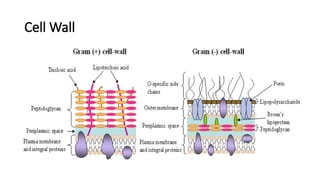
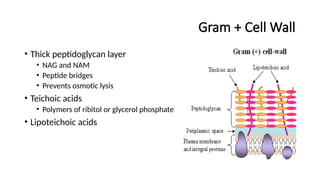
Gram + CellWall
• Thick peptidoglycan layer
• NAG and NAM
• Peptide bridges
• Prevents osmotic lysis
• Teichoic acids
• Polymers of ribitol or glycerol phosphate
• Lipoteichoic acids
11.
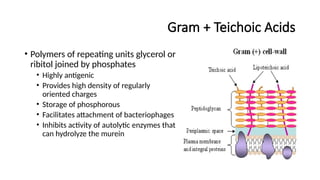
Gram + TeichoicAcids
• Polymers of repeating units glycerol or
ribitol joined by phosphates
• Highly antigenic
• Provides high density of regularly
oriented charges
• Storage of phosphorous
• Facilitates attachment of bacteriophages
• Inhibits activity of autolytic enzymes that
can hydrolyze the murein
12.
Gram + TeichoicAcids
Function WTA LTA
Anchoring
Structural Support Cell wall ____________ Cell wall ______________
Ion Regulation Maintains metal ion homeostasis
Immune Recognition Highly antigenic, triggers HIR Also antigenic/ immunogenic
Bacteriophage Attachment Recognized by bacteriophages Less involved
Autolysin Regulation Prevents ____________________ Controls _____________________
Virulence Factor Important for pathogenic adhesion
and immune evasion
contributory
13.
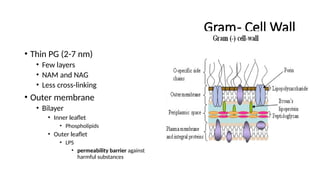
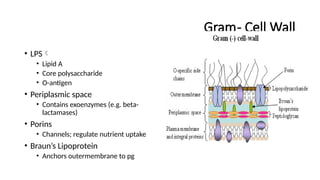
Gram- Cell Wall
•Thin PG (2-7 nm)
• Few layers
• NAM and NAG
• Less cross-linking
• Outer membrane
• Bilayer
• Inner leaflet
• Phospholipids
• Outer leaflet
• LPS
• permeability barrier against
harmful substances
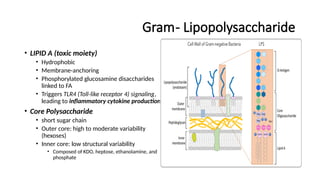
Gram- Lipopolysaccharide
• LIPIDA (toxic moiety)
• Hydrophobic
• Membrane-anchoring
• Phosphorylated glucosamine disaccharides
linked to FA
• Triggers TLR4 (Toll-like receptor 4) signaling,
leading to inflammatory cytokine production
• Core Polysaccharide
• short sugar chain
• Outer core: high to moderate variability
(hexoses)
• Inner core: low structural variability
• Composed of KDO, heptose, ethanolamine, and
phosphate
16.
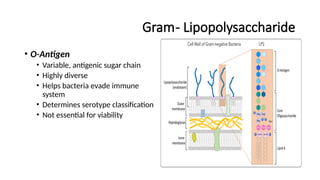
Gram- Lipopolysaccharide
• O-Antigen
•Variable, antigenic sugar chain
• Highly diverse
• Helps bacteria evade immune
system
• Determines serotype classification
• Not essential for viability
17.
Lipopolysaccharide Synthesis
Part Synthesis
LipidA Synthesized from UDP-NAG
Made in the CM by peripheral membrane proteins
Core Synthesized on Lipid A
Outer Core: each sugar is added one at a time to Lipid A
O-antigen Synthesized on a separate lipid carrier (undecaprenyl-P)
completed oligosaccharide chain is transferred to lipid A
core
Substances vs PeptidoglycanSynthesis
Substance Effect on PG Synthesis
Phosphonomycin Prevents synthesis of UDP-NAM from UDP-NAG
Cycloserine Inhibition of pentapeptide formation
Bacitracin Inhibits lysine incorporation into the PG; prevents
dephosphorylation of the carrier lipid
Vancomycin,
Tunicamycin,
Ristocetin
Inhibits translocation step of the PG synthesis
Penicillin Prevents cross-linking
25.
Mechanism of Action(MOA) of β-Lactam
Antibiotics
• Penicillin: weak penetration in the periplasmic space of Gram neg
• Inhibition of PBPs (Transpeptidases)
• Penicillin mimics D-Ala-D-Ala, binds irreversibly to PBPs, and blocks cross-
linking of peptidoglycan.
• Weakened Peptidoglycan Structure
• PG layer becomes unstable, making Gram-positive bacteria especially
vulnerable.
• Autolysin Activation → Cell Lysis
26.
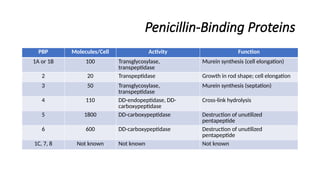
Penicillin-Binding Proteins
PBP Molecules/CellActivity Function
1A or 1B 100 Transglycosylase,
transpeptidase
Murein synthesis (cell elongation)
2 20 Transpeptidase Growth in rod shape; cell elongation
3 50 Transglycosylase,
transpeptidase
Murein synthesis (septation)
4 110 DD-endopeptidase, DD-
carboxypeptidase
Cross-link hydrolysis
5 1800 DD-carboxypeptidase Destruction of unutilized
pentapeptide
6 600 DD-carboxypeptidase Destruction of unutilized
pentapeptide
1C, 7, 8 Not known Not known Not known
27.
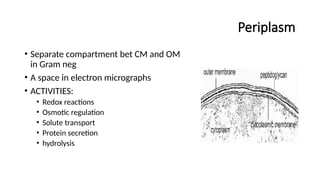
Periplasm
• Separate compartmentbet CM and OM
in Gram neg
• A space in electron micrographs
• ACTIVITIES:
• Redox reactions
• Osmotic regulation
• Solute transport
• Protein secretion
• hydrolysis
28.
Periplasm
Component Function
Oligosaccharides Involvedin osmotic regulation
Solute-Binding
Proteins
Bind to solutes and deliver them to
specific transporters in the CM
Cytochrome c Involved in ETC and redox rxn
Hydrolytic enzymes Degrade nutrients to smaller molecules
that can be transported across the CM
Detoxifying agents e.g. B-lactamase
TonB proteins Required for the uptake of solutes (iron
siderophores, vit B12) that do not
diffuse through the porin
29.
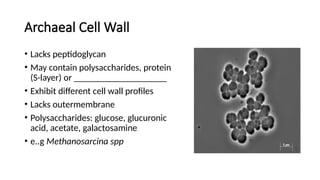
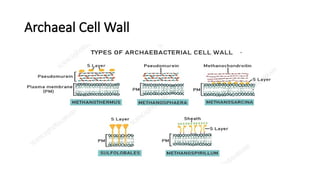
Archaeal Cell Wall
•Lacks peptidoglycan
• May contain polysaccharides, protein
(S-layer) or ____________________
• Exhibit different cell wall profiles
• Lacks outermembrane
• Polysaccharides: glucose, glucuronic
acid, acetate, galactosamine
• e..g Methanosarcina spp
Archaeal Cell Wall-S-Layer
• S-Layer (Surface Layer)
• Made of protein or glycoprotein subunits arranged in a paracrystalline
pattern.
• Forms a rigid, protective outer shell attached directly to the plasma
membrane.
• Example: Sulfolobus (thermoacidophile).
• Structure:
• Single-layer lattice of hexagonal, tetragonal, or linear symmetry.
• Pores allow nutrient exchange while blocking large molecules.
• Provides structural integrity and environmental resistance.
32.
Archaeal Cell Wall-Pseudomurein
• Pseudomurein (Pseudopeptidoglycan)
• Found in some methanogenic archaea (Methanobacterium,
Methanothermus).
Similar to bacterial peptidoglycan, but with key differences:
• N-acetyltalosaminuronic acid (NAT) instead of N-acetylmuramic acid (NAM).
• β-1,3 glycosidic bonds instead of β-1,4 (resistant to lysozyme).
• Peptide cross-links use L-amino acids, not D-amino acids like bacteria.
• Structure:
• Repeating units of NAT and NAG (N-acetylglucosamine) linked via β-1,3
bonds.
• Cross-linked peptide bridges provide strength.
33.
Mollicutes
• Mycoplasma andSpiroplasma
• Mostly parasitic and pathogenic
• Cell wall-less
• Gram-neg appearance
• pleiomorphic
• Have internal protein cytoskeleton
• Some contain unusually tough cytoplasmic membrane due to the
presence of sterols
#11 Explanation of Teichoic Acids
✅ Polymers of Repeating Units (Glycerol or Ribitol) Joined by Phosphates
Teichoic acids are long-chain polymers composed of either glycerol phosphate or ribitol phosphate.
These phosphate groups provide a negative charge, making the bacterial cell surface highly anionic.
✅ Highly Antigenic
Teichoic acids are highly immunogenic, meaning they can elicit strong immune responses in the host.
This antigenicity plays a role in host-pathogen interactions and bacterial virulence.
✅ Anchors __ to __
Wall teichoic acids (WTAs) are anchored to the peptidoglycan layer.
Lipoteichoic acids (LTAs) are anchored to the cytoplasmic membrane.
✅ Provides High Density of Regularly Oriented Charges
Due to their phosphate-rich structure, teichoic acids contribute to the electrostatic properties of the bacterial surface.
This influences interactions with metal ions, host cells, and antimicrobial peptides.
✅ Storage of Phosphorus
Since teichoic acids are phosphate-containing polymers, they can act as a reservoir of phosphorus when needed.
This is useful for bacterial survival under conditions of phosphorus limitation.
✅ Facilitates Attachment of Bacteriophages
Some bacteriophages (viruses that infect bacteria) recognize and bind to teichoic acids during infection.
This helps bacteriophages attach to and infect Gram-positive bacteria.
✅ Inhibits Activity of Autolytic Enzymes That Can Hydrolyze the Murein
Autolysins are enzymes that degrade peptidoglycan and regulate bacterial cell wall turnover.
Teichoic acids inhibit autolysins, preventing premature cell wall breakdown.
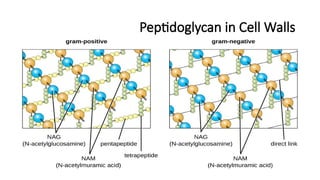
#18 Direction Type of Bond Function
X-Direction (Horizontal) β-1,4 Glycosidic Bond (NAG-NAM) Forms backbone, attacked by lysozyme
Y-Direction (Vertical) Peptide Bonds (NAM-NAM cross-links) Provides strength, targeted by β-lactam antibiotics
Basic Structure of the Pentapeptide
The pentapeptide attached to NAM typically follows this pattern:
🔹 L-Ala – D-Glu – meso-DAP (or L-Lys) – D-Ala – D-Ala
This sequence varies between Gram-positive and Gram-negative bacteria but usually includes:
L-Alanine (L-Ala)
D-Glutamic acid (D-Glu)
A diamino acid (meso-diaminopimelic acid (DAP) in Gram-negative, L-Lysine (L-Lys) in Gram-positive)
Two terminal D-Alanine (D-Ala – D-Ala)
The last D-Ala is lost during cross-linking, reducing it to a tetrapeptide in mature peptidoglycan.
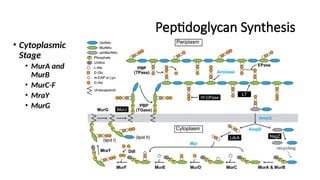
#20 Cytoplasmic Stage:
MurA & MurB: Convert UDP-NAG into UDP-NAM.
MurC-F: Sequentially add amino acids to form the pentapeptide.
MraY: Links UDP-NAM-pentapeptide to undecaprenol (C55-P), forming Lipid I.
MurG: Adds NAG, forming Lipid II.
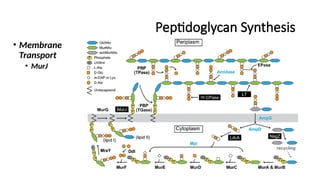
Membrane Transport:
MurJ (Flippase): Transports Lipid II across the membrane.
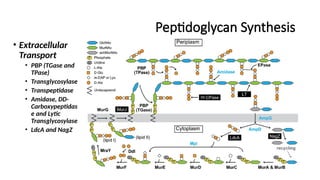
Extracellular Stage (Periplasmic Remodeling & Cross-Linking):
PBP (TGase & TPase):
Transglycosylase (TGase) polymerizes the NAG-NAM backbone.
Transpeptidase (TPase) catalyzes cross-linking between pentapeptides.
Amidase, DD-Carboxypeptidase (DD-CPase), and Lytic Transglycosylase (LT):
Modify the peptidoglycan mesh for controlled degradation and remodeling.
LdcA & NagZ: Recycle old peptidoglycan fragments.
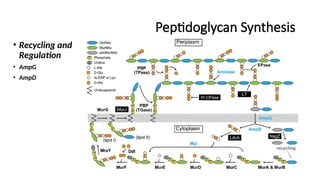
Recycling & Regulation:
AmpG & AmpD: Regulate peptidoglycan turnover and response to β-lactam antibiotics.
#21 Cytoplasmic Stage:
MurA & MurB: Convert UDP-NAG into UDP-NAM.
MurC-F: Sequentially add amino acids to form the pentapeptide.
MraY: Links UDP-NAM-pentapeptide to undecaprenol (C55-P), forming Lipid I.
MurG: Adds NAG, forming Lipid II.
Membrane Transport:
MurJ (Flippase): Transports Lipid II across the membrane.
Extracellular Stage (Periplasmic Remodeling & Cross-Linking):
PBP (TGase & TPase):
Transglycosylase (TGase) polymerizes the NAG-NAM backbone.
Transpeptidase (TPase) catalyzes cross-linking between pentapeptides.
Amidase, DD-Carboxypeptidase (DD-CPase), and Lytic Transglycosylase (LT):
Modify the peptidoglycan mesh for controlled degradation and remodeling.
LdcA & NagZ: Recycle old peptidoglycan fragments.
Recycling & Regulation:
AmpG & AmpD: Regulate peptidoglycan turnover and response to β-lactam antibiotics.
#22 Cytoplasmic Stage:
MurA & MurB: Convert UDP-NAG into UDP-NAM.
MurC-F: Sequentially add amino acids to form the pentapeptide.
MraY: Links UDP-NAM-pentapeptide to undecaprenol (C55-P), forming Lipid I.
MurG: Adds NAG, forming Lipid II.
Membrane Transport:
MurJ (Flippase): Transports Lipid II across the membrane.
Extracellular Stage (Periplasmic Remodeling & Cross-Linking):
PBP (TGase & TPase):
Transglycosylase (TGase) polymerizes the NAG-NAM backbone.
Transpeptidase (TPase) catalyzes cross-linking between pentapeptides.
Amidase, DD-Carboxypeptidase (DD-CPase), and Lytic Transglycosylase (LT):
Modify the peptidoglycan mesh for controlled degradation and remodeling.
LdcA & NagZ: Recycle old peptidoglycan fragments.
Recycling & Regulation:
AmpG & AmpD: Regulate peptidoglycan turnover and response to β-lactam antibiotics.
#23 Cytoplasmic Stage:
MurA & MurB: Convert UDP-NAG into UDP-NAM.
MurC-F: Sequentially add amino acids to form the pentapeptide.
MraY: Links UDP-NAM-pentapeptide to undecaprenol (C55-P), forming Lipid I.
MurG: Adds NAG, forming Lipid II.
Membrane Transport:
MurJ (Flippase): Transports Lipid II across the membrane.
Extracellular Stage (Periplasmic Remodeling & Cross-Linking):
PBP (TGase & TPase):
Transglycosylase (TGase) polymerizes the NAG-NAM backbone.
Transpeptidase (TPase) catalyzes cross-linking between pentapeptides.
Amidase, DD-Carboxypeptidase (DD-CPase), and Lytic Transglycosylase (LT):
Modify the peptidoglycan mesh for controlled degradation and remodeling.
LdcA & NagZ: Recycle old peptidoglycan fragments.
Recycling & Regulation:
AmpG & AmpD: Regulate peptidoglycan turnover and response to β-lactam antibiotics.
#25 1. Inhibition of Transpeptidase Activity (PBPs)
Penicillin binds to penicillin-binding proteins (PBPs), which function as transpeptidases.
PBPs normally cross-link the peptidoglycan peptide chains by forming covalent bonds between D-Ala-D-Ala of one PG unit and meso-DAP (or Lys) of another.
Penicillin mimics the D-Ala-D-Ala structure, irreversibly binding to PBPs and blocking cross-link formation.
2. Weakening of Peptidoglycan Structure
Without proper cross-linking, the PG layer becomes unstable and fragile.
This is especially fatal for Gram-positive bacteria, which rely on thick peptidoglycan for cell integrity.
In Gram-negative bacteria, β-lactams need to penetrate the outer membrane via porins to reach PBPs.
3. Activation of Autolysins → Cell Lysis
Many bacteria naturally produce autolysins that remodel peptidoglycan during cell growth.
Normally, PBPs counterbalance autolysins by continuously repairing the PG network.
When β-lactams block PBPs, autolysins continue to degrade PG, leading to osmotic lysis.
#27 Key Activities of the Periplasm
Redox Reactions
Facilitates oxidation and reduction processes.
Contains cytochromes and enzymes like Dsb proteins, which promote disulfide bond formation.
Osmotic Regulation
Maintains cell stability under changing osmotic conditions.
Contains osmoregulatory proteins (e.g., OsmY, MDOs) that adjust periplasmic composition based on external environment.
Solute Transport
Houses periplasmic binding proteins (PBPs) that aid in active transport via ABC transporters.
Acts as a buffer zone where molecules transition between the environment and cytoplasm.
Protein Secretion
Participates in Type II secretion system (T2SS) and Twin-arginine translocation (Tat) pathway to transport folded proteins.
Chaperones like SurA assist in outer membrane protein (OMP) folding.
Hydrolysis & Enzymatic Breakdown
Enzymes like β-lactamases degrade antibiotics, contributing to resistance.
Proteases (e.g., DegP) eliminate misfolded proteins.
Hydrolases break down complex molecules before transport into the cytoplasm.
#34 1. Carbon Linkage of Lipids
Bacteria & Eukarya: Lipids in their cell membranes have ester linkages.
Archaea: Use ether linkages instead.
Significance: Ether bonds are more chemically stable than ester bonds, allowing archaea to thrive in extreme environments (e.g., high temperatures, acidity, or salinity).
2. Phosphate Backbone of Lipids
Bacteria & Eukarya: Use glycerol-3-phosphate as the backbone for their membrane lipids.
Archaea: Use glycerol-1-phosphate instead.
Significance: The difference in lipid backbones reflects a fundamental divergence in membrane chemistry, supporting the hypothesis that archaea and bacteria evolved separately from a common ancestor.
3. Metabolism
Bacteria & Archaea: Have a bacterial-like metabolism, meaning they can perform diverse metabolic processes, including anaerobic respiration, fermentation, and nitrogen fixation.
Eukarya: Metabolism is distinct and includes specialized organelles (mitochondria and chloroplasts) for energy production.
4. Presence of a Nucleus
Bacteria & Archaea: No nucleus; their DNA is free-floating in the cytoplasm.
Eukarya: Has a nucleus, where DNA is enclosed by a nuclear membrane.
Significance: The presence of a nucleus is a major feature distinguishing eukaryotic cells from prokaryotic cells (Bacteria and Archaea).
5. Presence of Organelles
Bacteria & Archaea: No organelles.
Eukarya: Contains organelles such as mitochondria, endoplasmic reticulum, and Golgi apparatus.
Significance: Organelles allow compartmentalization of cellular functions, increasing efficiency.
6. Presence of Spliceosomal Introns (Non-coding DNA Sequences)
Bacteria & Archaea: No introns in their genes.
Eukarya: Has introns, which are removed during RNA processing.
Significance: Introns allow for gene regulation and alternative splicing, increasing genetic complexity in eukaryotes.
7. Presence of Telomeres
Bacteria & Archaea: No telomeres, as they have circular chromosomes.
Eukarya: Has telomeres, which protect the ends of linear chromosomes from degradation.
Significance: Telomeres prevent DNA loss during replication, a key feature in eukaryotic cell division and aging.
8. Chromosome Shape
Bacteria: Mostly circular chromosomes.
Archaea: Circular chromosomes.
Eukarya: Linear chromosomes.
Significance: Circular chromosomes are more stable and commonly found in prokaryotes, whereas linear chromosomes require telomeres for maintenance in eukaryotes.
9. DNA Replication
Bacteria: Uses bacterial-type replication enzymes.
Archaea: Uses eukaryotic-like replication enzymes.
Eukarya: Uses eukaryotic replication mechanisms.
Significance: This similarity between Archaea and Eukarya supports the hypothesis that Eukarya evolved from an archaeal ancestor.
10. Transcription (RNA Synthesis)
Bacteria: Uses bacterial transcription machinery.
Archaea: Uses eukaryotic-like transcription machinery.
Eukarya: Uses eukaryotic transcription.
Significance: Archaea and Eukarya share similarities in transcription enzymes, further indicating a close evolutionary relationship.
11. Translation (Protein Synthesis)
Bacteria: Uses bacterial ribosomes for translation.
Archaea: Uses eukaryotic-like ribosomes for translation.
Eukarya: Uses eukaryotic ribosomes.
Significance: Archaea's translation system resembles Eukarya’s more than Bacteria’s, reinforcing their evolutionary link.