Differential connectivity in neoplastic coexpression networks (BITS2014, Rome)
•
0 likes•306 views

Recommended

Recommended
More Related Content
What's hot
What's hot (20)
Viewers also liked
Viewers also liked (14)
Similar to Differential connectivity in neoplastic coexpression networks (BITS2014, Rome)
Similar to Differential connectivity in neoplastic coexpression networks (BITS2014, Rome) (20)
More from Roberto Anglani
More from Roberto Anglani (11)
Differential connectivity in neoplastic coexpression networks (BITS2014, Rome)
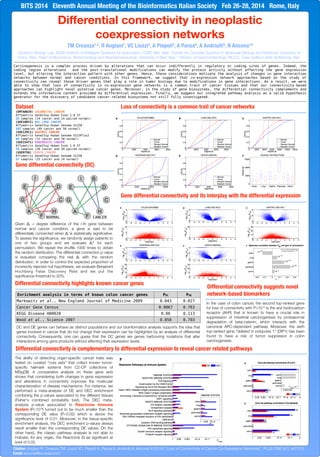
- 1. Differential connectivity in neoplastic coexpression networks TM Creanza1,2, R Anglani1, VC Liuzzi3, A Piepoli4, A Panza4, A Andriulli4, N Ancona1# 1Systems Biology Lab, ISSIA Institute of Intelligent Systems for Automation, CNR, Bari, Italy; 2Center for Complex Systems in Molecular Biology and Medicine, University of Torino, Italy; 3Dept of Bioscience, Biotechnology and Biopharmaceutical, University of Bari, Italy; 4 Division of Gastroenterology, IRCCS, Casa Sollievo della Sofferenza, Italy BITS 2014 Eleventh Annual Meeting of the Bioinformatics Italian Society Feb 26-28, 2014 Rome, Italy Carcinogenesis is a complex process driven by alterations that can occur indifferently in regulatory or coding sites of genes. Indeed, the coding region alterations and the post-translational modifications can modify the protein activity without affecting the gene expression level, but altering the interaction pattern with other genes. Hence, these considerations motivate the analysis of changes in gene interaction networks between normal and cancer conditions. In this framework, we suggest that co-expression network approaches based on the study of connectivity can reveal those driver genes that play a role in tumor biology due to modifications in gene interactions. As a result, we were able to show that loss of connectivity in co-expression gene networks is a common trait of cancer tissues and that our connectivity-based approaches can highlight novel putative cancer genes. Moreover, in the study of gene biosystems, the differential connectivity complements and extends the informative content provided by differential expression. Finally, we suggest our integrated pathway analysis as a valid hypothesis generator for the discovery of candidate cancer-related biosystems not still fully investigated. Loss of connectivity is a common trait of cancer networks Gene differential connectivity (DC) Differential connectivity suggests novel network-based biomarkers Differential connectivity is complementary to differential expression to reveal cancer related pathways Differential connectivity highlights known cancer genes Gene differential connectivity and its interplay with the differential expression Dataset (EMTAB829) COLORECTAL CANCER Affymetrix GeneChip Human Exon 1.0 ST 28 samples (14 cancer and 14 paired normal) (GSE10072) NSC LUNG CANCER Affymetrix GeneChip Human Genome U133A 107 samples (49 cancer and 58 normal) (GSE13911) GASTRIC CANCER Affymetrix GeneChip Human Genome U133Plus2 69 samples (31 cancer and 38 normal) (GSE15471) PANCREATIC CANCER Affymetrix GeneChip Human Exon 1.0 ST 78 samples (39 cancer and 39 paired normal) (GSE9750) CERVIX CANCER Affymetrix GeneChip Human Genome U133A 57 samples (33 cancer and 24 normal) Enrichment analysis in terms of known colon cancer genes PDC PDE Markowitz et al., New England Journal of Medicine 2009 0.043 0.027 Cancer Gene Census 0.0067 0.763 KEGG Disease H00020 0.06 0.113 Wood et al., Science 2007 0.058 0.703 Given Δi = degree difference of the i-th gene between normal and cancer condition, a gene is said to be differentially connected when Δi is statistically significative. To assess the significance, we randomly assign patients to one of two groups and we evaluate Δi* for each permutation. We repeat the shuffle 1000 times to obtain the random distribution. The differential connection p-value is evauated comparing the real Δi with the random distribution. In order to control the expected proportion of incorrectly rejected null hypotheses, we evaluate Benjamini Hochberg False Discovery Rate and we put the significance threshold to 20%. The ability of detecting organ-specific cancer traits was tested on curated “core sets” that collect known tumor- specific hallmark systems from C2-CP collections of MSigDB. A comparative analysis on these gene sets shows that considering both changes in gene expression and alterations in connectivity improves the molecular characterization of disease mechanisms. For instance, we performed a meta-analysis of DE and DEC enrichment combining the p-values associated to the different tissues (Fisher's combined probability test). The DEC meta- analysis p-value associated to Reactome Immune System (P<10-9) turned out to be much smaller than the corresponding DE value (P=0.02) which is above the significance level of 0.01. Moreover, in the tissue-specific enrichment analysis, the DEC enrichment p-values always result smaller than the corresponding DE values. On the other hand, the classic pathway analysis is not able to indicate, for any organ, the Reactome IS as significant at level of 0.05. In the case of colon cancer, the second top-ranked gene for loss of connectivity with P<10-3 is the aryl hydrocarbon receptor (AhR) that is known to have a crucial role in suppression of intestinal carcinogenesis by proteasomal degradation of beta-catenin, which interacts with the canonical APC-dependent pathway. Moreover, the sixth top-ranked gene “deleted in polyposis 1” (DIP1) has been found to have a role of tumor suppressor in colon carcinogenesis. DC and DE genes can behave as distinct populations and our bioinformatics analysis supports the idea that genes involved in cancer that do not change their expression can be highlighted by an analysis of differential connectivity. Consequently, one can guess that the DC genes are genes harbouring mutations that alter interactions among gene products without affecting their expression levels. NORMAL CANCER Citation: Anglani R, Creanza TM, Liuzzi VC, Piepoli A, Panza A, Andriulli A, Ancona N (2014) “Loss of Connectivity in Cancer Co-Expression Networks”. PLoS ONE 9(1): e87075. Email: ancona@ba.issia.cnr.it