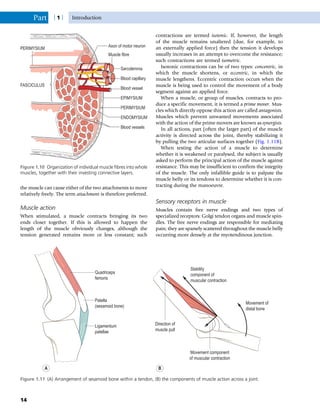
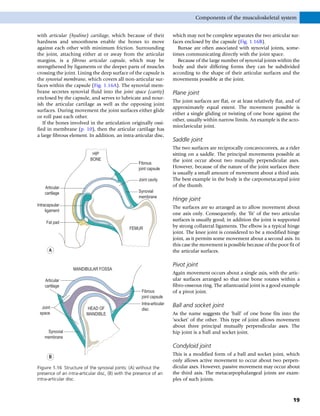
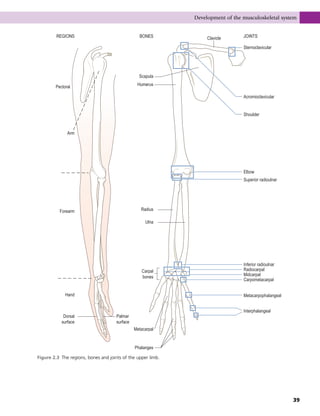
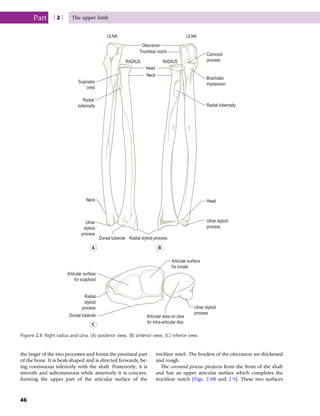
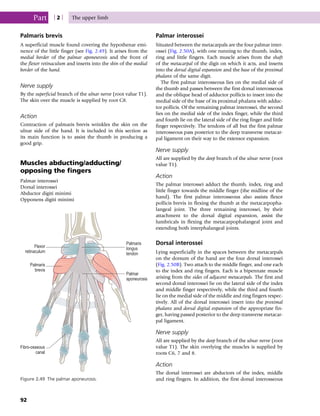
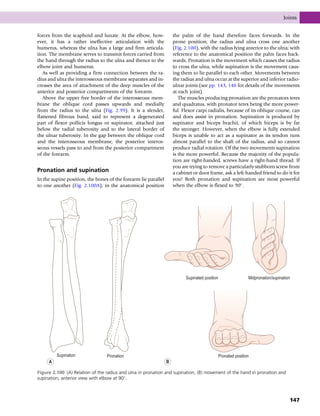
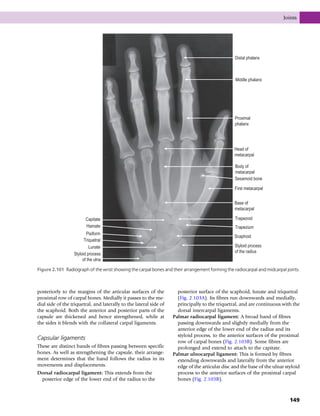
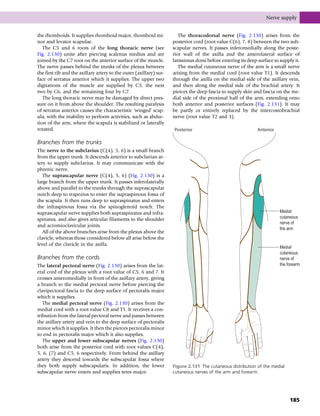
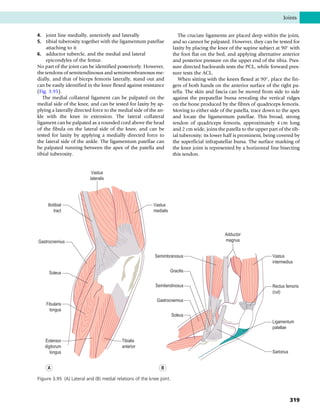
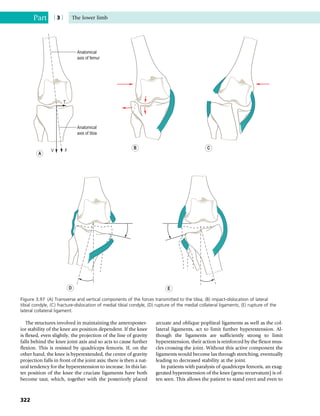
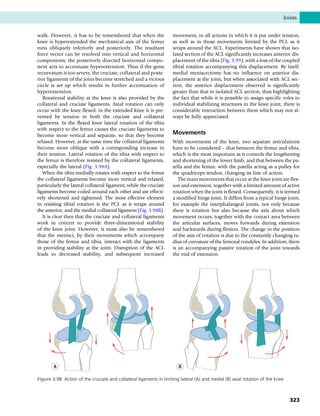
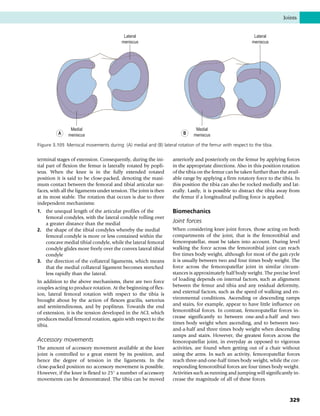
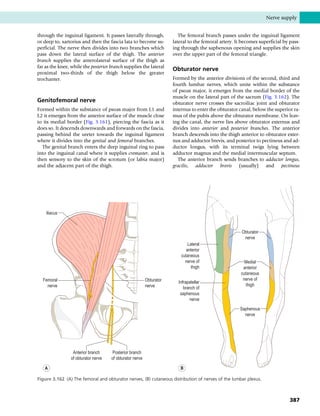
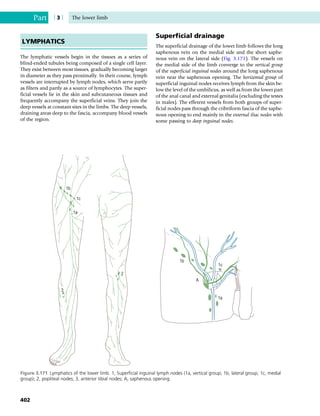
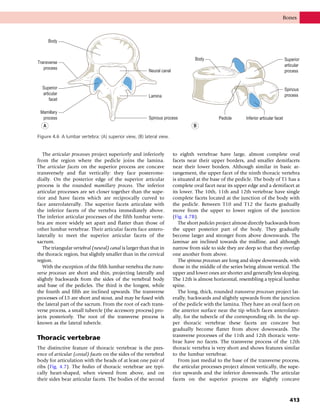
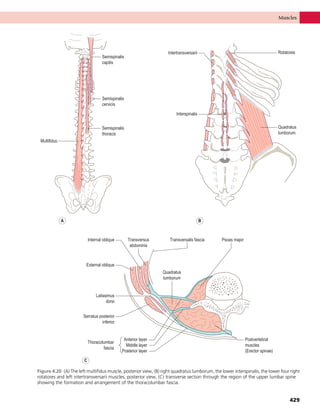
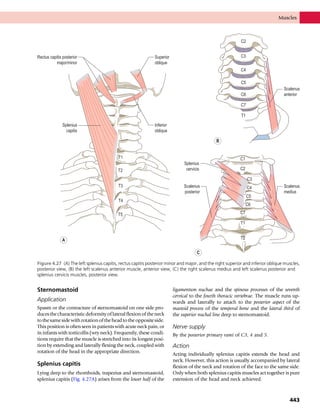
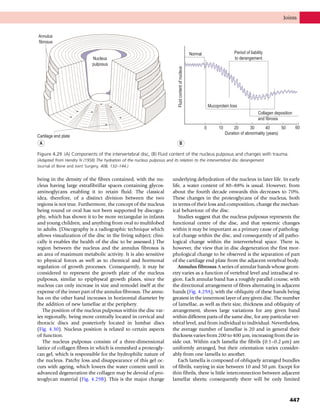
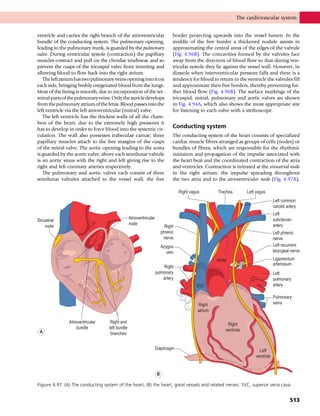
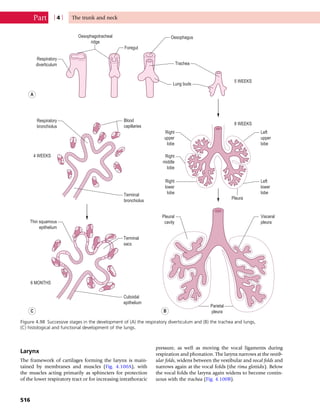
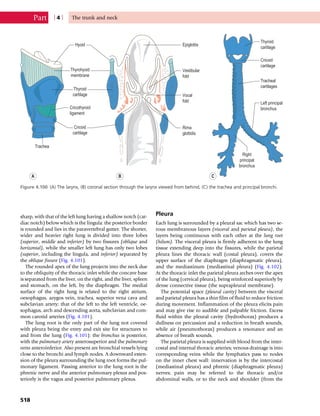
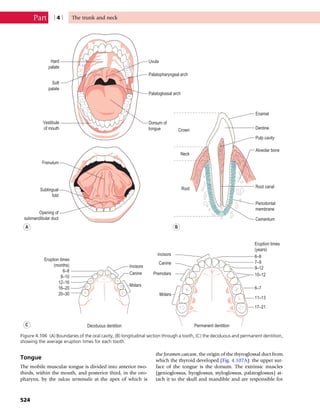
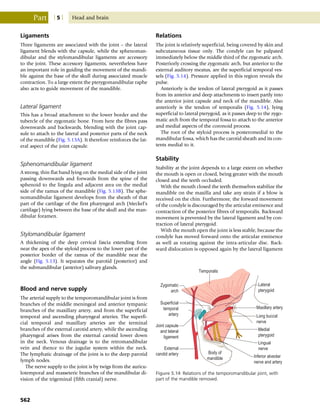
This document provides information about the sixth edition of the book "Anatomy and Human Movement: Structure and Function". It lists the commissioning editor, development editor, project manager, designer, illustrators, and publishers involved in producing the sixth edition. It also briefly describes the book's focus on describing musculoskeletal structures as a living dynamic system to aid understanding of human movement, rather than just as seen in dissection. The preface emphasizes the book's detailed illustrations of joints and biomechanics to explain function and potential failures. The goal is to fill a gap for students learning anatomy on living subjects to determine diagnoses and treatments.