The interaction of Nrf2 and Glyoxalase I in response to lipid loading in Hepa...
HHMI Poster Final
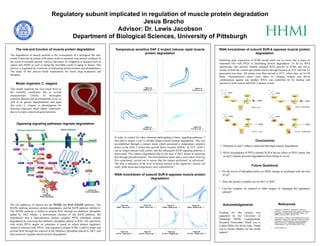
1. Regulatory subunit implicated in regulation of muscle protein degradation
Jesus Bracho
Advisor: Dr. Lewis Jacobson
Department of Biological Sciences, University of Pittsburgh
Acknowledgements
Conclusions
Future Questions
References
Opposing signaling pathways regulate degradation
The role and function of muscle protein degradation
1. Mutation in daf-2 induces rapid and thorough muscle degradation.
2. RNAi knockdown of PP2A subunit SUR-6 has no effect in PD55 strain, but
in daf-2 mutant prevents degradation from being as severe.
• Do the levels of phosphorylation on MEK change as predicted with the loss
of raf?
• Does the protein complex act on AKT or Raf?
• Can the complex be induced to other targets by changing the regulatory
subunit?
Funding for this project was
supported by the University of
Pittsburgh HHMI Undergraduate
Research Fellowship. Thank you to
Caitlin Miller for all the help. Thank
you to Jordan Sanders for the moral
support.
The degradation of muscle proteins is the consequence of a biological fail safe,
meant to provide an animal with amino acids to maintain vital protein synthesis in
the event of external stresses, such as starvation. It’s triggered in diseases such as
cancer and AIDS, as well as being the inevitable result of aging or disuse. This
process is regulated by a network of interacting protein kinases and phosphatases.
The study of this process holds implications for future drug treatments and
therapies.
Model organism C. elegans
This model organism has seen much favor in
the scientific community due to several
characteristics. Chiefly, its thoroughly
explored genome and developmental cycle, as
well as its genetic manipulability and rapid
life cycle. C. elegans is advantageous for
drawing conjecture about higher eukaryotes,
due to its many conserved gene functions.
The two pathways of interest are the MAPK and DAF-2/IGFR pathways. The
MAPK pathway promotes protein degradation, and the IGFR pathway inhibits it.
The MAPK pathway is halted at protein RAF through an inhibitory phosphate
added by AKT kinase, a downstream element of the IGFR pathway. We
hypothesize that a heterotrimeric protein complex PP2A stimulates protein
degradation by removing the inhibitory phosphate placed on RAF. The specificity
with which PP2A targets its substrates is based on which distinct regulatory
subunit it interacts with. PP2A, with regulatory subunit SUR6, is able to target and
activate RAF through the removal of the inhibitory phosphate placed by AKT, and
thus positively regulate muscle protein degradation.
Temperature sensitive DAF-2 mutant induces rapid muscle
protein degradation
RNAi knockdown of subunit SUR-6 opposes muscle protein
degradation
Figure 1c.
PD55 at 25°C
In order to control for other elements participating in these signaling pathways, I
first had to ensure a way to reliably induce muscle protein degradation. This was
accomplished through a mutant strain which possessed a temperature sensitive
defect in the DAF-2 insulin-like growth factor receptor (IGFR). At 25°C, DAF-2
can no longer interact with insulin, and the subsequent IGFR signaling pathway is
inactivated. This induces degradation due to the lack of AKT kinase to inactivate
Raf through phosphorylation. The histochemical stains above were taken from my
first experiment, carried out to ensure that the mutant performed ‘as advertised’.
The blue is indicative of the level of protein present in the organisms’ muscle cell
walls. Both strain and temperature were controlled for.
RNAi knockdown of subunit SUR-6 opposes muscle protein
degradation
Figure 2a.
PD55 + Null at 25°C
Figure 2b.
PD55 + Null at 25°C
Figure 2c.
PD55 + SUR6 at 25°C
Figure 2d.
PD55 + SUR6 at 25°C
Figure 2e.
PJ1741 + Null at 25°C
Figure 2f.
PJ1741 + Null at 25°C
Figure 2g.
PJ1741 + SUR6 at 25°C
Figure 2h.
PJ1741 + SUR6 at 25°C
Inhibiting gene expression of SUR6 would allow me to verify that it plays an
important role with PP2A in facilitating protein degradation. To do so, RNA
interference was utilized. Double stranded RNA specific to SUR6 was fed to
worms of both the control and mutant strains through bacteria at 16°C until the F1
generation was born. All strains were then moved to 25°C, where they sat for 48
hours. Histochemical stains were taken to compare mutant and RNAi
combinations against one another. RNAi was controlled for by feeding null
bacteria to both control and DAF-2 mutant worms.
DAF-2
Insulin
AKT Raf
FGFR
Autophagic
Degradation
Inhibitory
Phosphates
P
SUR-6
LET-92
PP2A
DAF-2
Insulin
AKT Raf
FGFR
Autophagic
Degradation
Inhibitory
Phosphates
P
SUR-6
LET-92
PP2A
DAF-2
Insulin
AKT Raf
FGFR
Autophagic
Degradation
Inhibitory
Phosphates
P
SUR-6
LET-92
PP2A
DAF-2
Insulin
AKT Raf
FGFR
Autophagic
Degradation
Inhibitory
Phosphates
P
SUR-6
LET-92
PP2A
PD55 + Null PD55 + SUR6 RNAi
PJ1741 + SUR6 RNAiPJ1741 + Null
Figure 1b.
PJ1741 at 16°C
Figure 1a.
PD55 at 16°C
Figure 1d.
PJ1741 at 25°C
1. Szewczyk N., Peterson B., Barmada S., Parkinson L.,
Jacobson L. “Opposed growth factor signals control protein
degradation in muscle of Caenorhabditis elegans.” The
EMBO Journal 26 (2007): 935-943. Print.
2. Padmanabhan, Srivatsan, Arnab Mukhopadhyay, Sri Devi
Narasimhan, Gregory Tesz, Michael P. Czech, and Heidi A.
Tissenbaum. "A PP2A Regulatory Subunit Regulates C.
Elegans Insulin/IGF-1 Signaling by Modulating AKT-1
Phosphorylation." Cell 136.5 (2009): 939-51. Print.
3. www.wormbase.org