1. S10 Oral Presentations / Growth Hormone & IGF Research 22S1 (2014) S5–S24
bGH mice revealed poorer memory performance, compared to
WT. GHA mice performed better and consistently during both the
acquisition and probe trials.
Conclusions: QPCR and Western analysis of the genes/proteins of
interest in the brain of GHA & their WT littermates and assessing
changes in the neurochemistry in these mice, will offer a better
understanding of neuroendocrinological aspect of GH/IGF-1 axis
and may identify potential therapeutic targets for neuropatho-
logical conditions.
OR3-2
GH enhances spine density in amygdalar neurons
B. Gisabella, J. Brophy, K.A. Goosens. McGovern Institute for Brain
Research, Massachusetts Institute of Technology, Cambridge, United
States
Introduction: Growth hormone (GH) exerts trophic effects in
many tissues throughout the body. Although GH is released into
the bloodstream by the pituitary gland, it is also synthesized
within limbic brain areas such as the amygdala, a brain region
that regulates fear memory. Despite this observation, little is
known about the effects of GH within the adult brain.
We previously reported a link between GH synthesized in the
amygdala and stress-related enhancement of fear memory.
Chronic stress in rodents increases GH levels in the amygdala and
overexpression of GH in amygdala neurons of unstressed rodents
mimics the fear-enhancing effect of chronic stress. However, it is
not clear how GH acts in the amygdala to enhance fear memory.
One possibility is that GH exerts neurotrophic effects.
Methods: To explore the hypothesis that GH produces neuro-
trophic effects in the adult brain, we used an adeno-associated
viral vector to overexpress either GH with green florescent protein
(GFP) or GFP alone in the basolateral complex of the amygdala
(BLA) in rats. Dendritic spine density was quantified by combin-
ing confocal imaging with three-dimensional dendritic analysis.
Results: We found that GH overexpression dramatically enhanced
the density of dendritic spines in the BLA (primary branches,
p=0.0002, secondary branches, p=0.0003).
Conclusion: This suggests that GH potently promotes dendritic
spinogenesis in neurons, illuminating a novel role for GH in the
adult brain, and provides a potential mechanism by which chronic
stress, which enhances GH in the amygdala, could contribute to
stress-induced alterations in amygdala morphology and function.
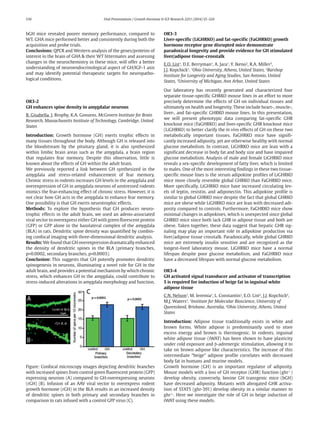
Figure: Confocal microscopy images depicting dendritic branches
with increased spines from control green fluorescent protein (GFP)
expressing neurons (A) compared to GH-overexpressing neurons
(rGH) (B). Infusion of an AAV viral vector to overexpress rodent
growth hormone (rGH) in the BLA results in an increased density
of dendritic spines in both primary and secondary branches in
comparison to rats infused with a control GFP virus (C).
OR3-3
Liver-specific (LiGHRKO) and fat-specific (FaGHRKO) growth
hormone receptor gene disrupted mice demonstrate
paradoxical longevity and provide evidence for GH stimulated
liver/adipose tissue-crosstalk
E.O. List1
, D.E. Berryman1
, A. Jara1
, Y. Ikeno2
, R.A. Miller3
,
J.J. Kopchick1
. 1
Ohio University, Athens, United States, 2
Barshop
Institute for Longevity and Aging Studies, San Antonio, United
States, 3
University of Michigan, Ann Arbor, United States
Our laboratory has recently generated and characterized four
separate tissue-specific GHRKO mouse lines in an effort to more
precisely determine the effects of GH on individual tissues and
ultimately on health and longevity. These include heart-, muscle-,
liver-, and fat-specific GHRKO mouse lines. In this presentation,
we will present phenotypic data comparing fat-specific GHR
knockout mice (FaGHRKO) and liver-specific GHR knockout mice
(LiGHRKO) to better clarify the in vivo effects of GH on these two
metabolically important tissues. FaGHRKO mice have signifi-
cantly increased adiposity, yet are otherwise healthy with normal
glucose metabolism. In contrast, LiGHRKO mice are lean with a
significant decrease in body fat and body size and have impaired
glucose metabolism. Analysis of male and female LiGHRKO mice
reveals a sex-specific development of fatty liver, which is limited
to males. One of the most interesting findings in these two tissue-
specific mouse lines is the serum adipokine profiles of LiGHRKO
mice more closely resemble global GHRKO than FaGHRKO mice.
More specifically, LiGHRKO mice have increased circulating lev-
els of leptin, resistin, and adiponectin. This adipokine profile is
similar to global GHRKO mice despite the fact that global GHRKO
mice are obese while LiGHRKO mice are lean with decreased adi-
posity compared to controls. Furthermore, FaGHRKO mice show
minimal changes in adipokines, which is unexpected since global
GHRKO mice since both lack GHR in adipose tissue and both are
obese. Taken together, these data suggest that hepatic GHR sig-
naling may play an important role in adipokine production via
liver/adipose tissue crosstalk. Paradoxically, while global GHRKO
mice are extremely insulin sensitive and are recognized as the
longest-lived laboratory mouse, LiGHRKO mice have a normal
lifespan despite poor glucose metabolism, and FaGHRKO mice
have a decreased lifespan with normal glucose metabolism.
OR3-4
GH activated signal transducer and activator of transcription
5 is required for induction of beige fat in inguinal white
adipose tissue
C.N. Nelson1
, M. Ieremia1
, L. Constantin1
, E.O. List2
, J.J. Kopchick2
,
M.J. Waters1
. 1
Institute for Molecular Bioscience, University of
Queensland, Brisbane, Australia, 2
Ohio University, Athens, United
States
Introduction: Adipose tissue traditionally exists in white and
brown forms. White adipose is predominantly used to store
excess energy and brown is thermogenic. In rodents, inguinal
white adipose tissue (iWAT) has been shown to have plasticity
under cold exposure and b-adrenergic stimulation, allowing it to
take on brown adipose like characteristics. The increase of this
intermediate “beige” adipose profile correlates with decreased
body fat in humans and murine models.
Growth hormone (GH) is an important regulator of adiposity.
Mouse models with a loss of GH receptor (GHR) function (ghr-/-
)
develop obesity, conversely, bovine GH transgenic mice (bGH)
have decreased adiposity. Mutants with abrogated GHR activa-
tion of STAT5 (ghr-391) develop obesity in a similar manner to
ghr-/-
. Here we investigate the role of GH in beige induction of
iWAT using these models.
2. Oral Presentations / Growth Hormone & IGF Research 22S1 (2014) S5–S24 S11
Method: Transcript and protein analysis of bGH, ghr-/-
, ghr-391
and iWAT determined the beige phenotype of these mouse
models. Plasma and tissue analysis was performed for key beige
inducer, FGF21. Beige cell induction via FGF21 infusion and
b3-adrenergic stimulation was tested in GHR mutants and their
wt littermates. Beige induction was measured by transcript and
protein analysis and histologically.
Results: The transcript profile of the iWAT revealed decreased
beige adipose markers in GHR mutants, but increased in bGH
mice. Proteins such as UCP1 and sub-units of the mitochondrial
oxidative phosphorylation complex are increased in bGH and
decreased in ghr-391. Despite low circulating and local FGF21 in
GHR mutants, FGF21 infusion failed to induce beige adipose inghr-
/-
and ghr-391 mice. b3-adrenergic stimulation was also ineffective.
Conclusions: GH is important in the development of beige fat.
Mice with a loss of GHR STAT5 activation are unable to induce beige
adipose in iWAT stores even with conventional inducers. Ghr-391
mice indicate STAT5 is critical for GH induction of beige cells.
OR3-5
NPY neurons as a critical hypothalamic node for the control
of GH release relative to food intake
L. Huang1
, H. Tan1
, M. Fogarty1
, R. Stark2
, Z. Andrews2
, J. Veldhuis3
,
H. Herzog4
, C. Chen1
, F. Steyn1
. 1
School of Biomedical Sciences,
University of Queensland, Brisbane, Australia, 2
Department of
Physiology, Monash University, Melbourne, Australia, 3
Department
of Medicine, Endocrine Research Unit, Mayo Clinic, Rochester,
United States, 4
Neuroscience Research Program, Garvan Institute of
Medical Research, Sydney, Australia
Neuropeptide-Y (NPY) expressing neurons are orexigenic neurons
that sense negative energy balance and engage neural mecha-
nisms to restore energy balance by increasing food intake and
decreasing energy expenditure. We previously documented the
suppression of pulsatile GH secretion in the fasting mouse, and
demonstrated that this occurs alongside a rise in hypothalamic
NPY and somatostatin mRNA expression. Given anticipated inter-
actions between NPY and somatostatin neurons, we proposed
that NPY neurons act through somatostatin neurons to suppress
GH release in the fasting mouse.
We confirmed interactions between somatostatin and NPY
expressing neurons in the mouse by demonstrating synaptic sites
between NPY fibres and somatostatin positive projections within
the periventricular nucleus. Using NPY-deficient (NPYKO) mice,
we demonstrate the complete recovery of pulsatile GH release
in fasting mice. Importantly, GH pulsatility in NPYKO mice did
not change under fed conditions, suggesting that NPY neurons
primarily participate in GH release during negative energy bal-
ance. NPY exerts its effects through multiple NPY-responsive
receptors (Y-receptors). The Y1 receptor (Y1R) is the dominant
postsynaptic receptor, whereas the Y2 receptor (Y2R) is mainly
expressed presynaptically on NPY neurons. We confirmed the
recovery of pulsatile GH release in fasting germ-line deleted Y1R
(Y1RKO) mice, whereas germ-line deletion of the Y2R (Y2RKO)
did not recover the fasting-induced suppression of GH release.
Rather, we observed a significant reduction in peak GH secretion
in ad libitum fed Y2RKO mice.
Observations confirm that NPY suppresses GH secretion in the
mouse through postsynaptic interactions with the Y1 receptor. As
NPY neurons do not participate in the regulation of GH pulsatility
under fed conditions, we propose that the Y2R contributes to GH
release independent of NPY. Collectively, observations confirm
that NPY neurons, acting through somatostatin neurons, are a
key sensory node integrating metabolic information between the
body and hypothalamic regulators of GH release.
OR3-6
Loss of acyl-ghrelin signalling in male ghrelin-O-acyltransferase
knock-out mice results in reduced pulsatile growth hormone
secretion and a derangement of GH pulse pattern
T.Y. Xie1
, S.T. Ngo1
, J.D. Veldhuis2
, P.L. Jeffery3
, L.K. Chopin3
,
M. Tschöp4
, V. Trolle5
, J. Epelbaum5
, F.J. Steyn1
, C. Chen1
. 1
School of
Biomedical Sciences, University of Queensland, Brisbane, Australia,
2
Department of Medicine, Endocrine Research Unit, Mayo School of
Graduate Medical Education, Clinical Translational Science Center,
Rochester, United States, 3
Ghrelin Research Group, Translational
Research Institute, Queensland University of Technology, Brisbane,
Australia, 4
Institute for Diabetes and Obesity, Helmholtz Zentrum
München, German Research Center for Environmental Health,
Neuherberg, Germany, 5
UMR-S 894 INSERM, Centre de Psychiatrie
et Neurosciences, Université Paris Descartes, Paris, France
Introduction: Ghrelin is a nutrient-sensing hormone primarily
secreted by oxyntic cells of the stomach and gastrointestinal tract.
Its biological activity is regulated by the acylation at serine-3
with 8-carbon fatty acid, catalysed by ghrelin-O-acyltransferase
(GOAT) enzyme. Exogenous acyl-ghrelin augments the release of
GH, however speculation remains whether endogenous ghrelin
directly contributes to GH secretion.
Methods: We assessed pulsatile GH secretion in 8-, 16- and
36-weeks old ad libitum fed male germ-line GOAT-/-
mice. Starting
at 0700 h, 36 sequential tail-tip whole blood samples (4 ml/sam-
ple) were collected over a 6-h period at 10-min intervals. Analysis
for GH was performed using an in-house mouse GH ELISA and
quantified by deconvolution analysis. Assessment of pulsatile
GH secretion relative to respective ages, epididymal fat mass and
circulating levels of leptin were performed by linear regression
and Spearman correlation coefficient analyses.
Results: Observations show a significant reduction in overall
GH secretion (total, pulsatile GH secretion and mean peak per
GH pulse) in GOAT-/-
mice at 8 and 16 weeks of age. We observed
an age-associated rise in body weight, epididymal fat mass and
circulating levels of leptin in both genotypes. This was observed
alongside an age-related decline in overall GH secretion. A rise in
the number of GH secretory events (pulse number) and approxi-
mate entropy (increased irregularity) was observed in GOAT-/-
mice regardless of age. This did not change relative to epididymal
fat weight or circulating levels of leptin.
Conclusions: Observations confirm that acyl-ghrelin mediates
peak GH release in male mice, independent of age and adiposity.
Moreover, as we demonstrate a rise in GH pulse frequency and a
derangement of pulse pattern in GOAT-/-
mice, we propose that
acyl-ghrelin may be essential for the maturation, development or
integration of hypothalamic or peripheral GH pulse generators.