
Recommended
More Related Content
What's hot
What's hot (17)
Viewers also liked
Similar to Reproductive senescence has negative effects on early egg development and embryonic viability in Drosophila-5
Similar to Reproductive senescence has negative effects on early egg development and embryonic viability in Drosophila-5 (20)
Reproductive senescence has negative effects on early egg development and embryonic viability in Drosophila-5
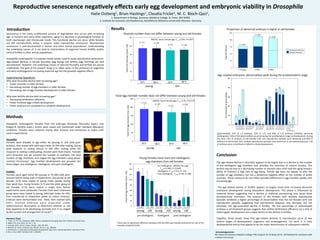
- 1. n-‐=11 0 20 40 60 80 100 120 140 160 0 10 20 30 40 50 Reproduc)ve senescence nega)vely effects early egg development and embryonic viability in Drosophila Halie Ostberg1, Brian Has3ngs1, Claudia Fricke2, M. C. Bloch-‐Qazi1, 1. Department of Biology, Gustavus Adolphus College, St. Peter, MN 56082 2. Ins3tute for Evolu3on and Biodiversity, WesSälische Wilhelms-‐Universität, Münster, Germany Conclusion The age-‐related decline in fecundity appears to be largely due to a decline in the number of pre-‐vitellogenic egg chambers and, possibly, the reten3on of mature oocytes. This decline may be due to a decreasing number of ovariole stem cells6 and the aging female’s ability to maintain a high rate of egg laying. Female age does not appear to alter the number of egg chambers, but has a borderline nega3ve effect on the number of ac3ve ovarioles. These measures do not reflect possible differences in egg chamber quality with increasing age. The age-‐related decline in fer3lity appears to largely result from increased abnormal embryonic development during blastoderm development. This phase is influenced by maternal factors sugges3ng that a decline in maternal provisioning may cause these developmental anomalies. The embryos of old females with mul3ple reproduc3ve episodes exhibited a higher percentage of abnormali3es than the old females with one reproduc3ve episode, sugges3ng that reproduc3ve diapause may decrease, but not eliminate, the age-‐associated decline in fer3lity. The low percentage of undeveloped embryos in all treatment groups suggests that neither fer3liza3on efficiency nor a lack of ini3al zygo3c development are a major factors in the decline in fer3lity. Together, these results show that age-‐related declines in reproduc3on occur at two dis3nct stages of development: oogenesis and embryogenesis. For each, it is early developmental events that appear to be the major determinants of subsequent viability. Acknowledgements: We thank the Gustavus Adolphus College FYRE program for funding and Dr. Jeff Dahlseid for assistance with confocal microscopy Senescence is the many mul3faceted process of degrada3on that occurs with increasing age. In humans and many other organisms, aging is a decrease in physiological func3on at both macroscopic and microscopic levels. This func3onal decline can occur while females are s3ll reproduc3vely ac3ve, a process called reproduc3ve senescence. Reproduc3ve senescence is well-‐documented in human and other animal popula3ons1. Understanding the underlying causes of it can lead to interven3ons to augment human fer3lity and/or control fer3lity in other animal popula3ons. Drosophila melanogaster is a popular model system used to study reproduc3ve senescence. Age-‐related declines in female fecundity (egg laying) and fer3lity (egg hatching) are well documented2. However, the underlying causes of reduced fecundity and fer3lity are poorly understood. The goal of the present study is to refine when in the processes of oogenesis and early embryogenesis increasing maternal age has the greatest nega3ve effects. Experimental Ques3ons Why does fecundity decline with increasing age? • Fewer ovarioles in older females • Decreasing number of egg chambers in older females • Decreasing rate of egg chamber development in older females Why does fer3lity decline with increasing age? • Decreasing fer3liza3on efficiency • Fewer fer3lized eggs ini3ate development • Fewer embryos are competent to complete development Results Pre-‐vitellogenic: ANOVA, fem age F1,44=11.080, P< 0.0005 Vitellogenic: F1,44=1.512, P=.226 Post-‐vitellogenic: F1,44=3.156, P=.084 Literature Cited 1. Finch, C.E. & T.B.L. Kirkwood. 2000. Chance, Development & Aging. New York: Oxford University Press. 2. Miller, P.B., et al. 2014. Fly. 8:3, 1-‐13. 3. Cumings, M.R., and R.C. King J. Morph., 128: 427-‐442. 4. Bownes, M. 1975 J. Embryol. Exp. Morph. Vol.33, 3, pp. 789-‐801. 5. Hartenstein, V. 1993 Atlas of Drosophila Development. New York: Cold Spring Harbor Laboratory Press. 6. Pan, L. et al. 2007. Cell Stem Cell 1, 458-‐469. ANOVA, factor female age, F1, 41=3.66, P=0.063 24 h 24 h 96 h 96 h young old n-‐=11 n-‐=10 n-‐=11 n-‐=10 Ovariole number does not differ between young and old females Age-‐related embryonic abnormali3es peak during the preblastoderm stage Methods Drosophila melanogaster females from the wild-‐type Dahomey (fecundity expts.) and Oregon-‐R (fer3lity expts.) strains were raised and maintained under standard laboratory condi3ons. Females were collected shortly arer eclosion and maintained as virgins un3l used in experiments. Fer)lity Females were aged either 8d (young) or 35-‐38d (old) post-‐ eclosion before being used in experiments. One group of old females (3-‐O) were mated to young males weekly during their adult lives. Young females (1-‐Y) and the other group of old females (1-‐O) were mated a single 3me before experiments were conducted. Females from each treatment group were mass mated to young, wild-‐type males for 24h, then transferred to oviposi3on plates to collect embryos. Embryos were dechorinated and fixed, then stained with DAPI. Stained embryos were examined under epiflourescence illumina3on to determine whether or not they appeared to be developing normally and, if so, staged by the number and arrangement of nuclei4,5. Fecundity Females were allowed to age either 4d (young) or 32d (old) post-‐ eclosion, then mated with wild-‐type males. At 24h arer ma3ng, during peak response to ma3ng s3muli, or 96h arer ma3ng, while the response to ma3ng is asenua3ng, females were flash-‐frozen. Females were dissected and we counted the number of ovarioles, the total number of egg chambers, and staged the egg chambers using phase-‐ contrast microscopy3. Egg chamber development was grouped into three stages: pre-‐vitellogenic, vitellogenic, and post-‐vitellogenic. 0 50 100 150 200 250 0 10 20 30 40 50 Approximately 12% of 1-‐Y embryos, 32% of 1-‐O, and 40% of 3-‐O embryos exhibited abnormal development. 65% of the abnormali3es occurred during the preblastoderm stage of development. During this 3me, 75% of embryos of old females with one reproduc3ve episode were abnormal and 84% of embryos of old females with mul3ple reproduc3ve episodes were abnormal. In all treatment groups, <3% of embryos were unfer3lized or failed to ini3ate development. Abnormal Preblastoderm Embryo Normal Preblastoderm Embryo Propor3on of abnormal embryos is higher in old females 0 50 100 150 200 250 n-‐=154 n-‐=225 n-‐=216 Total # Embryos 1-‐Young 1-‐Old 3-‐Old % of embryos at given phase Abnormal embryos Normal embryos 1-‐Y 1-‐O 3-‐O Preblastoderm ovaries n-‐=11 n-‐=10 n-‐=11 n-‐=10 young old 24 h 96 h 24 h 96 h 24 h 24 h 24 h 24 h 24 h 24 h 96 h 96 h 96 h 96 h 96 h 96 h young old old old young young pre-‐vitellogenic vitellogenic post-‐vitellogenic Total egg chamber number does not differ between young and old females ANOVA, factor female age, F1,41=1.91, P=0.174 Young females have more pre-‐vitellogenic egg chambers than old females There was no significant difference between 24h and 96h post-‐ma3ng treatments for any of the variables tested (P>0.05). Total # of Ovarioles (mean +SD) Total # of egg chambers (man +SD) Total # of egg chamber (mean +SD) Introduc)on 1-‐Y 1-‐O 3-‐O 1-‐Y 1-‐O 3-‐O 1-‐Y 1-‐O 3-‐O 1-‐Y 1-‐O 3-‐O 1-‐Y 1-‐O 3-‐O Blastoderm Gastrula3on Segmenta3on Dorsal Closure Cu3cle Specializa3on 13 48 79 12 6 3 67 6 9 2 42 31 58 90 79 23 5 16 Ovaries Egg chambers