1. Biochemical & protein interaction analysis of calcium-sensing
proteins CML 15 and CML 16 in Arabidopsis thaliana
Matt Watson
Supervisor: Dr. Wayne Snedden, Committee Members: Dr. Sharon Regan & Dr. Virginia Walker
METHOD: Protein purification & calcium-dependent
hydrophobic affinity chromatography
INTRODUCTION
METHOD: Western Blotting
OBJECTIVE 1: Generation of recombinant
CMLs for use in affinity chromatography.
v Recombinant CML 15 & CML16 were successfully purified in
a manner consistent with canonical Ca2+ sensors such as
CaM
v CML 16 has detectable expression in 10-day WT seedling
tissue from Arabidopsis thaliana Col-0 WT
v Evidence that CML 16 has putative binding targets within the
CD4-30 library, suggesting that a positive target has been
isolated from floral tissue. This target will need to be further
corroborated with further in vitro and in vivo methods.
This study has set the foundation for future study into the
downstream targets of CML 15 & 16. Future work into these
CMLs should focus on the development of affinity columns
using recombinant CMLs, as well as corroboration of putative
targets from the yeast 2 hybrid screen.
CONCLUSIONS
METHOD: Yeast 2 hybrid screen- use of a bait CML
vector to identify putative binding interactions with
a prey vector from a floral cDNA library
.
OBJECTIVE 3: Identify tissue-specific
expression patterns of CML 16.
RESULT: Conjugation of both recombinant CMLs to
phenyl-Sepharose reconfirmed their characteristics
as putative Ca2+ sensors & produced soluble
protein fractions for affinity chromatography.
RESULT: Western Blotting confirmed detection of
CML 16 in 10-day old seedling tissue. Evidence of
CML 16 expression provided the rationale for its use
in future tissue-specific assays.
RESULT: Putative binding interactions were observed
between CML 16 (pGBKT7 vector) and targets in the
CD4-30 floral library, suggesting a putative binding
target for CML 16 in floral tissue.
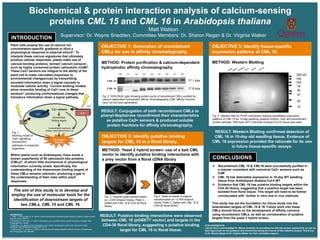
Fig. 2. SDS-PAGE gels showing protein purity of recombinant CMLs purified by
calcium-dependent hydrophobic affinity chromatography (CML affinity columns
have not yet been generated).
Fig. 5. Western Blot on PVDF membrane showing quantitative expression
patterns of CML 16 for 10-day seedling (positive control), root, and recombinant
protein samples. Wild-type (WT) Colombia Ecotype (Col-0) tissue was used.
Fig. 3. Original yeast transformation
on –LTAH dropout media, Plate C,
plated with CML 16 & CD4-30 floral
library.
OBJECTIVE 2: Identify putative binding
targets for CML 16 in a floral library.
Fig.4. Yeast re-streak of original
transformation on –LTAH dropout
media, Plate C, plated with CML 16 &
CD4-30 floral library.
Within plants such as Arabidopsis, there exists a
known superfamily of 50 calmodulin-like proteins
(CMLs)4, of which little biochemical or physiological
information currently exists. Specifically, the
understanding of the downstream binding targets of
these CMLs remains unknown, producing a gap in
the understanding of their roles within plant
responses.
The aim of this study is to develop and
employ the use of molecular tools for the
identification of downstream targets of
two CMLs, CML 15 and CML 16.
REFERENCES
1. Yang, T., & Poovaiah, B. W. (2003). Calcium/calmodulin-mediated signal network in plants. Trends in plant
science, 8(10), 505-512.
2. Snedden, W. & Fromm, H. (2001). Calmodulin as a versatile calcium signal transducer in plants. New
Phytologist 151, 35–66.
3. DeFalco, T.A., Bender, K.W. & Snedden, W.A. (2010). Breaking the code: Ca2+ sensors in plant
signaling. The Biochemical Journal 425, 27-40.
4. McCormack, E. & Braam, J. (2003). Calmodulins and related potential calcium sensors of Arabidopsis.
New Phytologist 159, 585-598.
Acknowledgments
I would like to acknowledge Dr. Wayne Snedden for providing me with this project opportunity, as well as
Deni Ogunrinde for her guidance and mentorship during the course of this research project. Thank you
to Dr. Sharon Regan & Dr. Virginia Walker for their committee support.
Fig. 1.
Working model of
Ca2+ signalling
transduction
pathways in eukaryotic
organisms.
Plant cells employ the use of calcium ion
concentration-specific gradients to illicit a
physiological response to external stimuli1. To
interpret these calcium signatures that ultimately
produce cellular responses, plants make use of
calcium-binding proteins, termed ‘calcium sensors’,
such as highly conserved protein calmodulin (CaM)2.
These Ca2+ sensors are integral to the ability of the
plant cell to make calculated responses to
environmental changes/cues by transmitting
encoded information down a signal cascade to
modulate cellular activity. Current working models
show reversible binding of Ca2+ ions to these
sensors3, producing conformational changes that
transduce information down a signal pathway.
17.1 kda
17.6 kda
10
20
37
50
75
100
150
250 kD