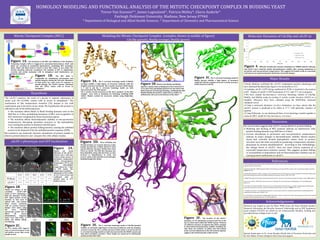
1. HOMOLOGY MODELING AND FUNCTIONAL ANALYSIS OF THE MITOTIC CHECKPOINT COMPLEX IN BUDDING YEAST
Trevor Van Eeuwen*^, James Luginsland^, Patricia Melloy*, Gloria Anderle^
Fairleigh Dickinson University, Madison, New Jersey 07940
* Department of Biological and Allied Health Sciences, ^ Department of Chemistry and Pharmaceutical Science
Mitotic Checkpoint Complex (MCC)
cdc20-1 phenotype and GFP localization
0
References
Acknowledgements
Major Results
Figure 1A. The formation of the MCC and inhibition of the Anaphase
Promoting Complex (APC) is triggered by unattached kinetochores. Mad1 and
Mad3 are recruited to unattached kinetochores, facilitating MCC formation.
Mad2, Mad3 and Bub3 bind to Cdc20 to form MCC which in turn binds APC
thereby blocking substrate recognition. The inability of APC to ubiquitinate
securins and cyclins maintains cells in metaphase until kinetochores are
properly attached.
Hypothesis
Mad1 Mad2
Mad3
Bub3
Cdc20
Cdc20
Mad3
Bub3
Mad2
MCC
Cdc20
binding
event
Cdc20
APC
Unattached
Kinetochore
Inactivated APC
Mad3
Bub3
Mad2
Figure 1B. The MCC (left) is
responsible for maintaining chromosome and
spindle integrity during the cell cycle. Mutations
in these genes can cause errors in response to
spindle damage. The Cdc20-1 mutant (obtained
from the ATCC) causes cells to arrest in
metaphase.1
The cdc20-1 mutation, the result of a glycine to arginine substitution at
amino acid 544 (G544R), causes cells to arrest in metaphase.2 The
mechanism of this temperature sensitive (Ts) mutant is not well
understood and is the focus of our study. We hypothesize the protein is
affected in one of the following ways:
The mutation affects Mad2 or Mad3 binding domains such as the
KEN Box or D Box, prohibiting formation of MCC and recognition of
APC substrates recognized by these functional regions.
The mutation affects thermodynamic stability at non-permissive
temperatures, disrupting secondary structure or the hydrophobic
core causing the protein to denature and fall apart.
The mutation affects protein folding kinetics, causing the unfolded
protein to be disposed of by the unfolded protein response (UPR).
Special thanks goes to Dr. Grant Murphy (Hecht lab) of Princeton University and
Dr. Eric Muller of Iona College for their time and support.
Research was funded in part by Pfizer PURE Grant, the Novo Nordisk Summer
Research Fellowship, the Novartis Research Scholarship and an NSF Equipment
grant number 0721251 for which we are immeasurably thankful. Funding also
provided Becton College Grant–in-Aid.
25oC 27oC
30oC 32oC 37oC
wt
cdc20-1
wt
cdc20-1
wt
cdc20-1
wt
cdc20-1
25oC 27oC 30oC 32oC 37oC
Wildtype +++ +++ +++ +++ +++
cdc20-1 +++ +++ ++ +/- -
DIC GFP DNA
Figure 3A. The S. cerevisiae homology model of Mad3p
(orange) exhibits a high degree of structural similarity with the
template structure (textured). The S. pombe structure (4AEZ chains
C, F and I) and the S. cerevisiae homology model are both
characterized by the TPR repeats.
N.B. S. pombe crystal structures of the three members of the MCC
complex were used as templates for homology modeling (PDB
4AEZ)4. Models verified for structural validity using Molprobity
server5.
Figure 3B. Shown is the Mad3p and Mad2p
interface. Amino acids on Mad2p (F134 E137 K140 E174
V175 and Y194) and Mad3p (Q18 N19 I21 K37 R40 V198
R202) network via hydrogen bonding , stabilizing the MCC.
Mad2p/Mad3p interaction is important as neither are
sufficient for cell cycle in the absence of the other6.
Figure 3D. Areas of Mad3p and
Cdc20p interface are of great interest as they
include the KEN Box of Mad3 (K30 E31 N32)
and the KEN Box Receptor (D260 D261 F262
Y263) on Cdc20. The KEN Box Receptor has
been identified, along with the D Box, as sites
involved in ubquitination of cyclin and
securin. amino acids typically act by hydrogen
bonding though side chains.
Figure 3F. The location of the cdc20-1
mutation, a G→R substitution at amino acid 544, at the
top of a beta sheet within the WD40 repeats. Pictured
are different rotamers of arginine that will be sampled
to determined thermodynamic stability including the
side chain out (yellow), in (blue) and self-cyclized
(red). Current simulations of the self-cyclized arginine
suggest a thermodynamically stable protein.
Figure 3E. The S. cerevisiae homology model of Cdc20p (purple)
exhibits an extremely high degree of structural similarity with the template
structure (textured). The S. pombe structure (4AEZ chains A, D and G) and the
S. cerevisiae homology model are both characterized by the WD40 repeats and
beta sheet secondary structure. These motifs are conserved in functionally
similar proteins like Cdh1.
Figure 3C. The S. cerevisiae homology model of
Mad2p (green) exhibits a high degree of structural
similarity with the template structure S. pombe structure
(4AEZ chains B, E and H). Serial dilutions confirmed the cdc20-1 temperature-sensitive phenotype and
have indicated the phenotype begins to be noticeable at 32 C.
Candidate cdc20-1-GFP (being confirmed by PCR) is localized to the nucleus
at RT. Studies of cdc20-1-GFP localization at 32 C and 37 C are in progress.
We have created Saccharomyces cerevisiae homology models of Cdc20p,
Mad2p, and Mad3p using an existing S. pombe crystal structure (4AEZ) as a
template. Structures have been validated using the MolProbity structural
validation server.
Using a molecular dynamics in-silico simulation, we have shown that the
cdc20-1 protein is predicted to be stable at 37 C with particular rotamers of
arginine.
Using PyRosetta, we are docking all three of our homology models together to
create an MCC model for Saccharomyces cerevisiae7.
We wanted to use molecular dynamic simulations of protein models to
observe protein behavior and compare this with cellular studies.
Figure 4. RMS plot of molecular dynamic simulations in AMBER indicate wild type
and cdc20-1 have relatively similar thermodynamic stability after running simulations at
permissive and non-permissive temperatures. This suggests all models reside in a potential
well and are not experiencing short-timeframe structural changes.
Molecular Dynamics of Cdc20p and cdc20-1p
Figure 2A.
Shown are serial
dilutions of the cdc20-1
mutant and a wild type
control strain grown at
permissive temperature
(25oC), non-permissive
temperature (37oC) and
three intermediate
temperatures for 7 days
on YPD media. The
temperature-sensitive
phenotype is first visible
at 32oC (red arrow) and
confirmed with no
growth seen at 37oC
(yellow arrow).
Figure 2C.
At 37oC, cdc20-1-GFP signal is
seen at several locations in the
cell (yellow arrow, blue arrow,
purple arrow).
Figure 2B.
Shown are images of the
candidate cdc20-1-GFP
strain at 25oC containing a
DNA dye (Hoechst 33342).
GFP signal (yellow arrow)
localizes to bud neck of
mitotic cells and nucleus
(blue arrow), consistent
with wt. protein location.3
Images were taken using
the Leica DM5500
microscopy system and an
ORCA ER camera
(Hamamatsu). Exposure
times were 5ms (DIC), 2s
(GFP) and 400ms (DNA),
2x2 binning.
1. Hartwell LH, Mortimer RK, Culotti J, Culotti M (1973). Genetic control of the cell division cycle in yeast. V. Genetic analysis of cdc
mutants. Genetics, 74:267-286.
2. Schott EJ, Hoyt MA (1998). Dominant alleles of Sacchromyces cerevisiae CDC20 reveal its role in promoting anaphase. Genetics
148(2):599-610.
3. Melloy PM, Holloway S (2004). Changes in the Localization of the Saccharomyces cerevisiae Anaphase-Promoting Complex Upon
Microtubule Depolymerization and Spindle Checkpoint Activation. Genetics, 167:1079-1094.
4. Chao WCH, Kulkarni K, Zhang Z, Kong EH, Barford D (2012). Structure of the mitotic checkpoint complex. Nature, 484: 208-214.
5. Chen et al. (2010). MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallographica
D66:12-21.
6. Tavorima PA, Burke DJ (1998). Cell cycle arrest in cdc20 mutants of Saccharomyces cerevisiae is independent of Ndc10p and
kinetochore function but requires a subset of spindle checkpoint genes. Genetics. 148(4): 1701–1713.
7. Chaudhury S, Lyskov S, Gray JJ (2010). PyRosetta: a script-based interface for implementing molecular modeling algorithms
using Rosetta. Bioinformatics., 26:689–691
8. Tan KP, Khares S, Varadarajan R, Madhusudhan MS (2014). Tspred: a web server for the rational design of temperature sensitive
mutants. Nucelic Acids Research. doi: 10.1093/nar.gku319.
Modeling the Mitotic Checkpoint Complex (complex shown in middle of figure)
Cdc20p (purple), Mad3p (orange), Mad2p (green)
Discussion
Our current data suggests that cdc20-1 causes defects in protein folding.
Modeling and docking of MCC proteins indicate no interference with
protein binding domains (e.g. KEN Box or D Box).
Current simulations at permissive and non-permissive temperatures
indicate no major changes in thermodynamic stability. Recent studies
found that centrally located hydrophobic amino acids in protein
structure were the most successful at producing a temperature sensitive
phenotype by protein destabilization8. According to this methodology,
the change found in cdc20-1 does not meet criteria expected of a
successful temperature-sensitive mutant. This suggests protein folding
or susceptibility to degradation and not thermodynamic stability may be
causing protein malfunction in cdc20-1.