Objectives:
•
* Recognize: -Self-MHC Restriction of T helper and Cytotoxic T Cells.
- Processing of Ag is required for recognition by T Cells.
* Explain professional and non- professional Ag presenting Cells.
* Discuss: -The Cytosolic Pathway (Endogenous Antigens ) and
assembly & stabilization of class I MHC molecules.
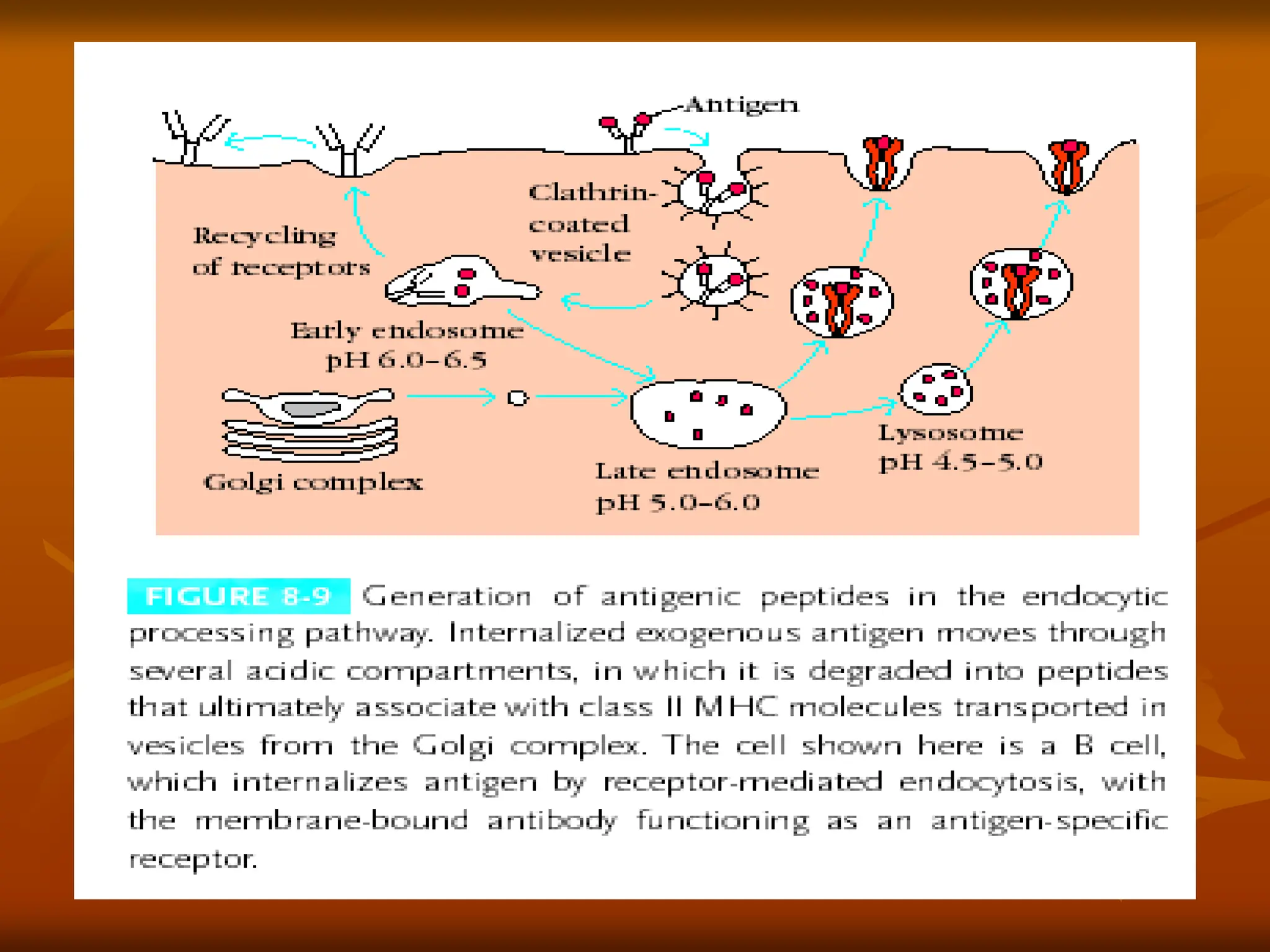
-The Endocytic Pathway (Exogenous Antigens) and
assembly of class II MHC molecules.
* Explain overview of endogenous and exogenous pathways for
processing antigen
•
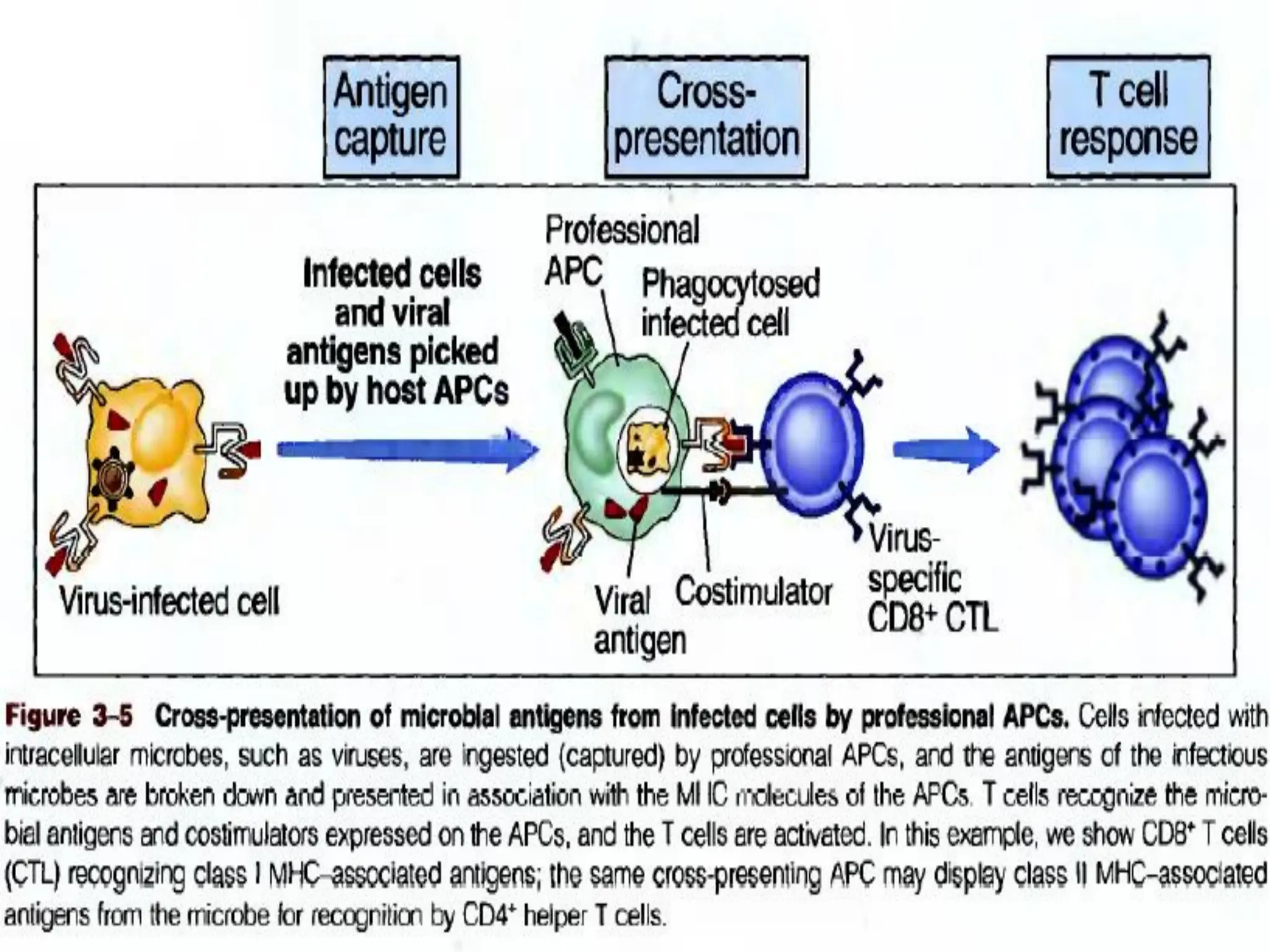
* Recogniz Cross presentation of Exogenous antigens.
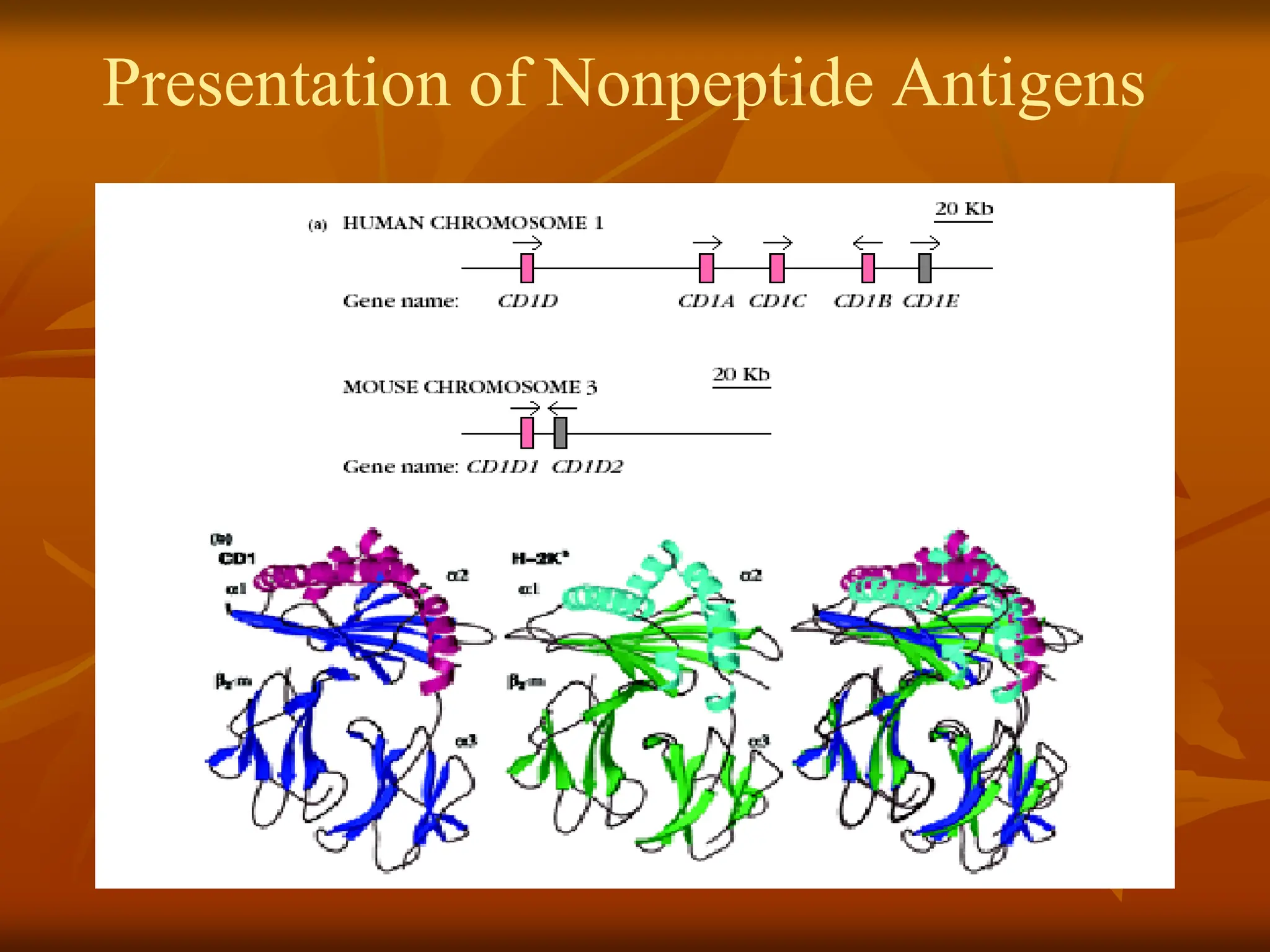
* Expain Presentation of Nonpeptide Antigens.
•
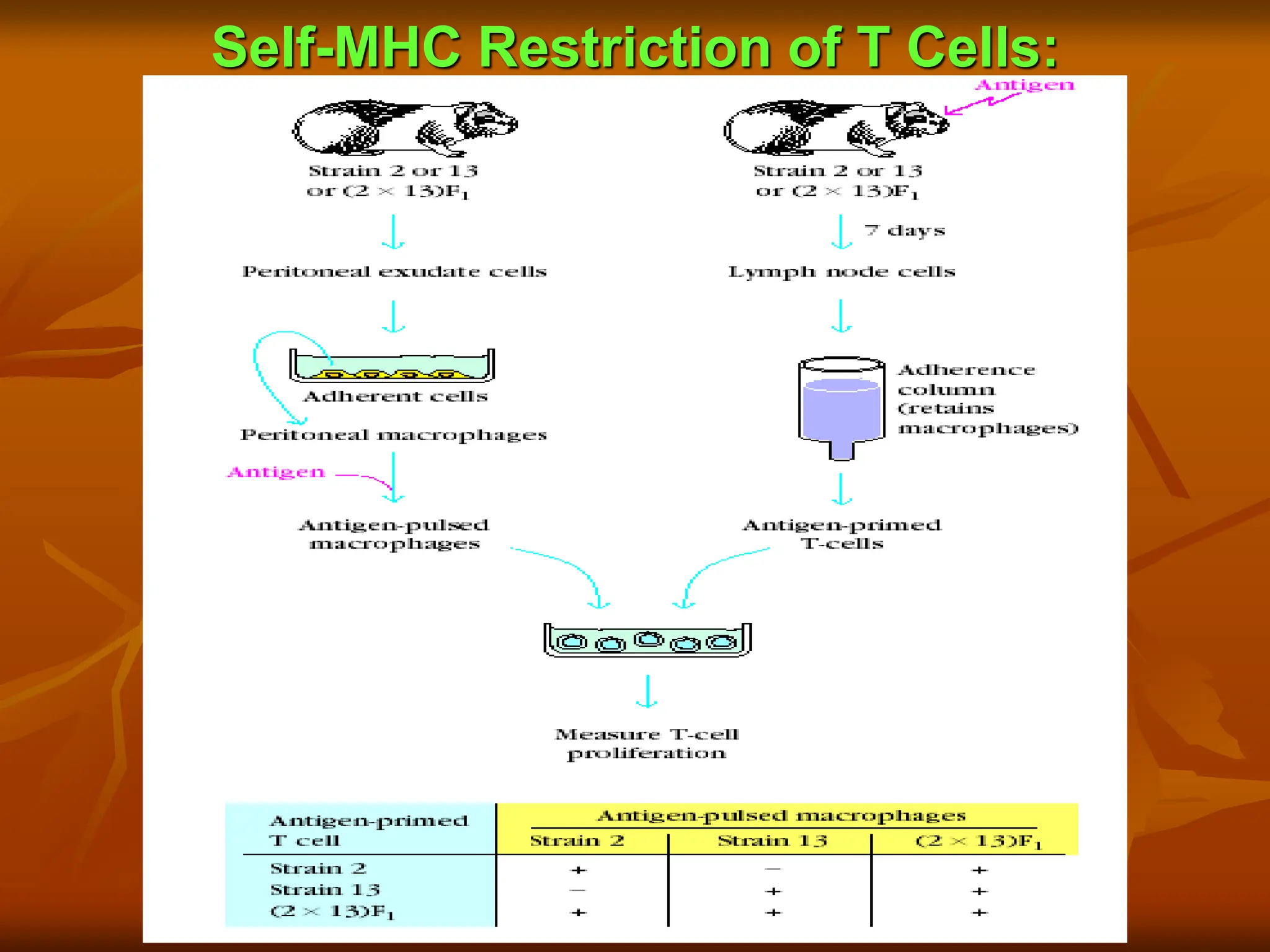
FIGURE . Experimentaldemonstration of self-MHC
restriction of TH cells. Peritoneal exudate cells from
strain 2, strain 13, or (2 X 13) F1 guinea pigs were
incubated in plastic petri dishes, allowing enrichment of
macrophages, which are adherent cells. The peritoneal
macrophages were then incubated with antigen. These
“antigen-pulsed” macrophages were incubated in vitro
with T cells from strain 2, strain 13, or (2 X 13) F1 guinea
pigs, and the degree of T-cell proliferation was assessed (+
vs -). The results indicated that TH cells could proliferate
only in response to antigen presented by macrophages
that shared MHC alleles.
7.
•
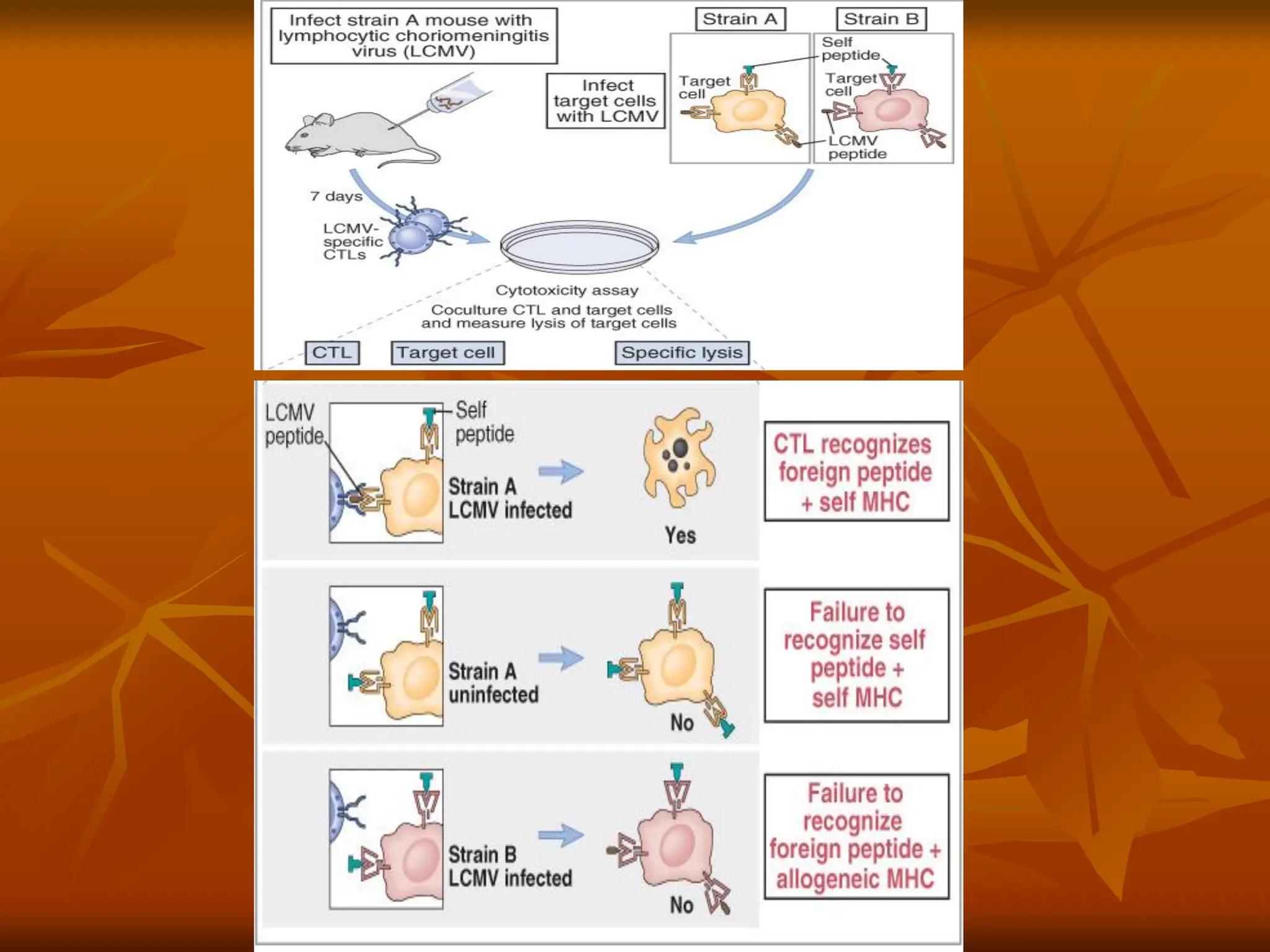
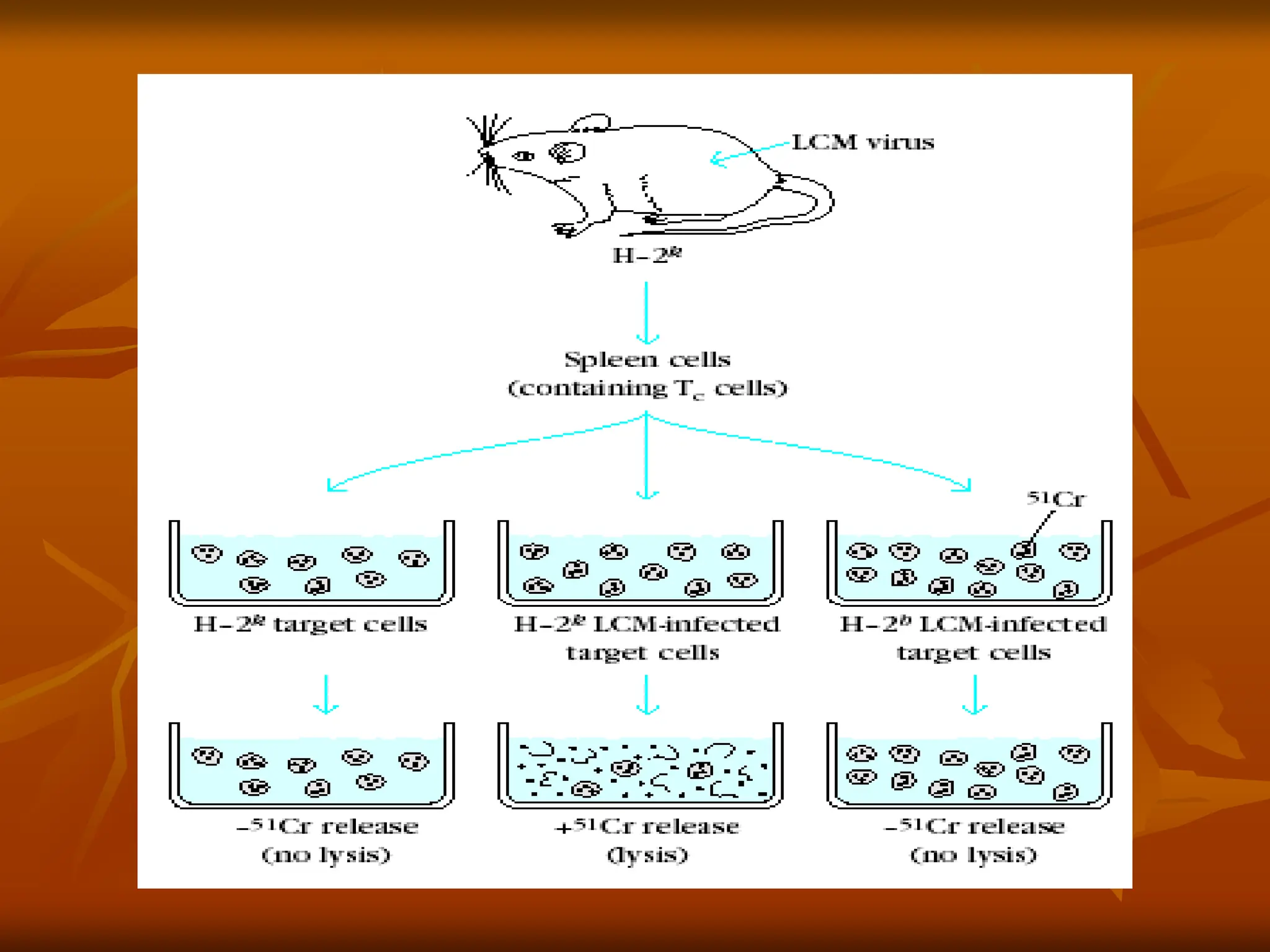
Classic experiment ofZinkernagel and Doherty
demonstrating that antigen recognition by TC cells
exhibits MHC restriction. H-2k mice were primed
with the lymphocytic choriomeningitis virus (LCMV) to
induce cytotoxic T lymphocytes (CTLs) specifi c for the
virus. Spleen cells from this LCMV-primed mouse were
then added to target cells of different H-2 haplotypes
that were intracellularly labeled with 51Cr (black dots)
and either infected or not with the LCMV. CTL-mediated
killing of the target cells, as measured by the release of
51Cr into the culture supernatant, occurred only if the
target cells were infected with LCMV and had the same
MHC haplotype as the CTLs.
•
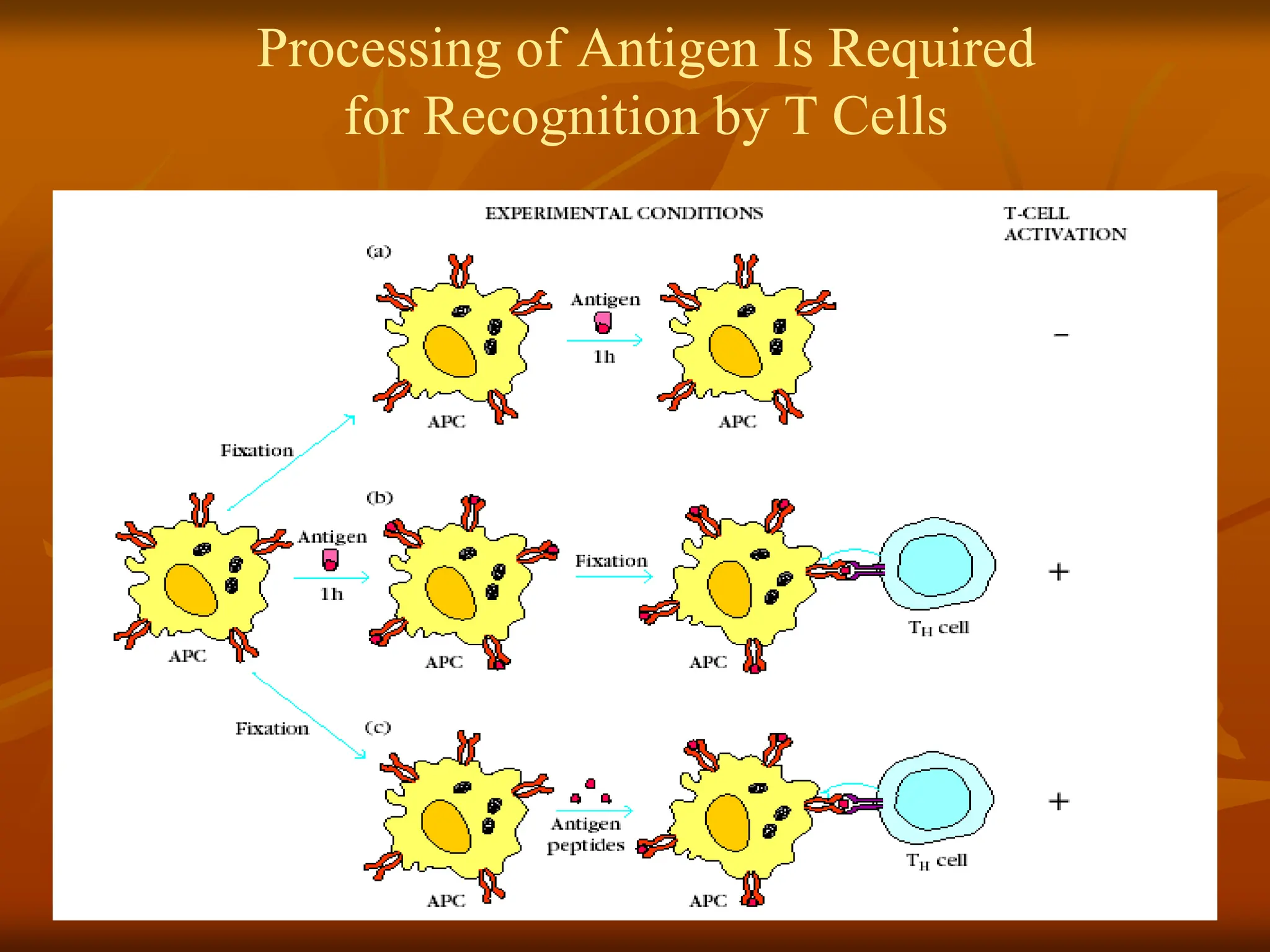
FIGURE - Experimentaldemonstration that antigen
processing is necessary for TH-cell activation. (a)
When presenting cells (APCs) are fixed before exposure
to antigen, they are unable to activate TH cells. (b) In
contrast, APCs fi xed at least 1 hour after antigen
exposure can activate TH cells. (This simplified figure
does not show costimulatory molecules needed for T-cell
activation.) (c) When APCs are fixed before antigen
exposure and incubated with peptide digests of the
antigen (rather than native antigen), they also can
activate TH cells. TH-cell activation is determined by
measuring a specifi c TH-cell response (e.g., cytokine
secretion).
12.
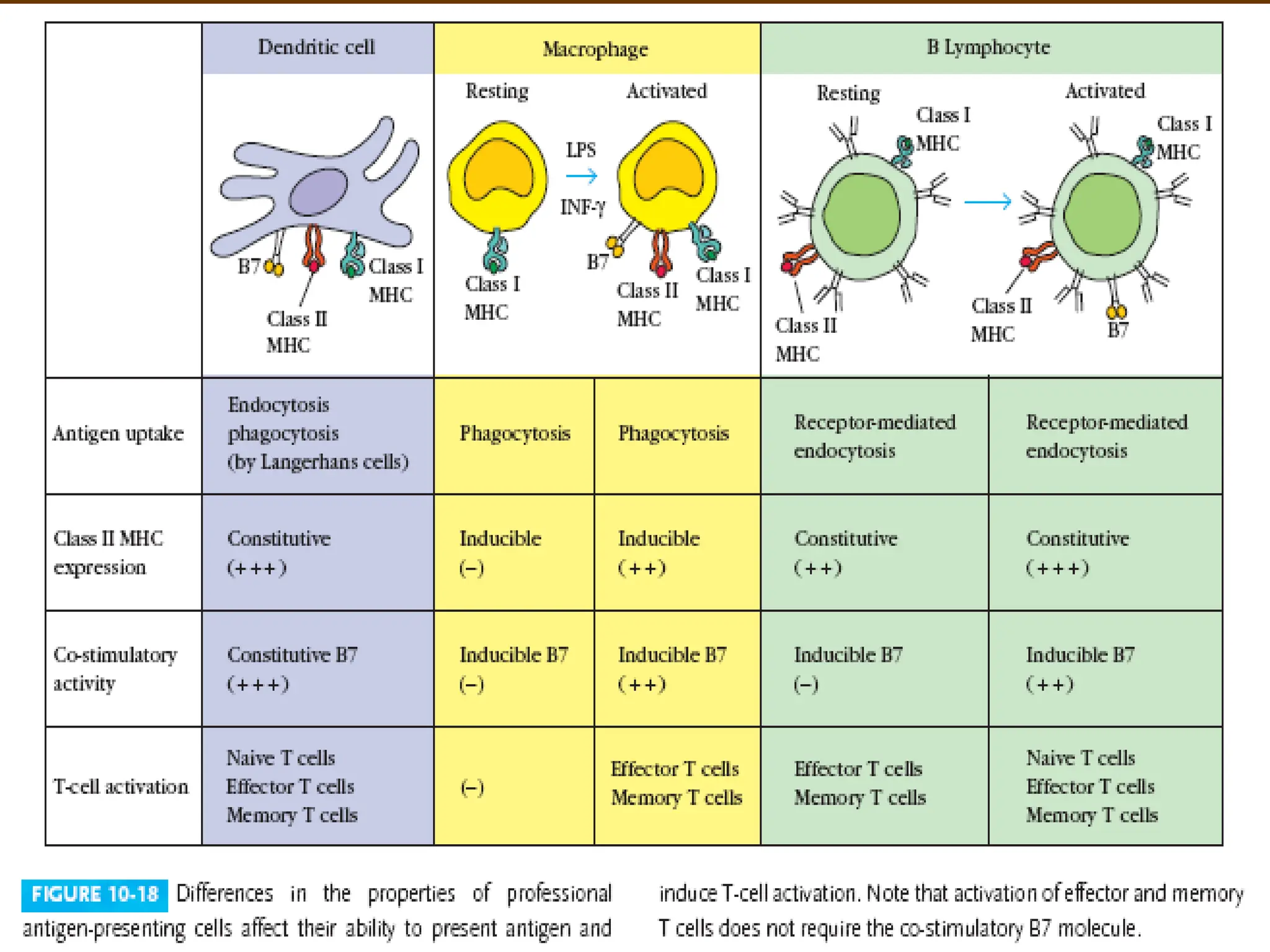
Most Cells CanPresent Antigen with
Class I MHC
Presentation with Class II MHC
Is Restricted to APCs
•
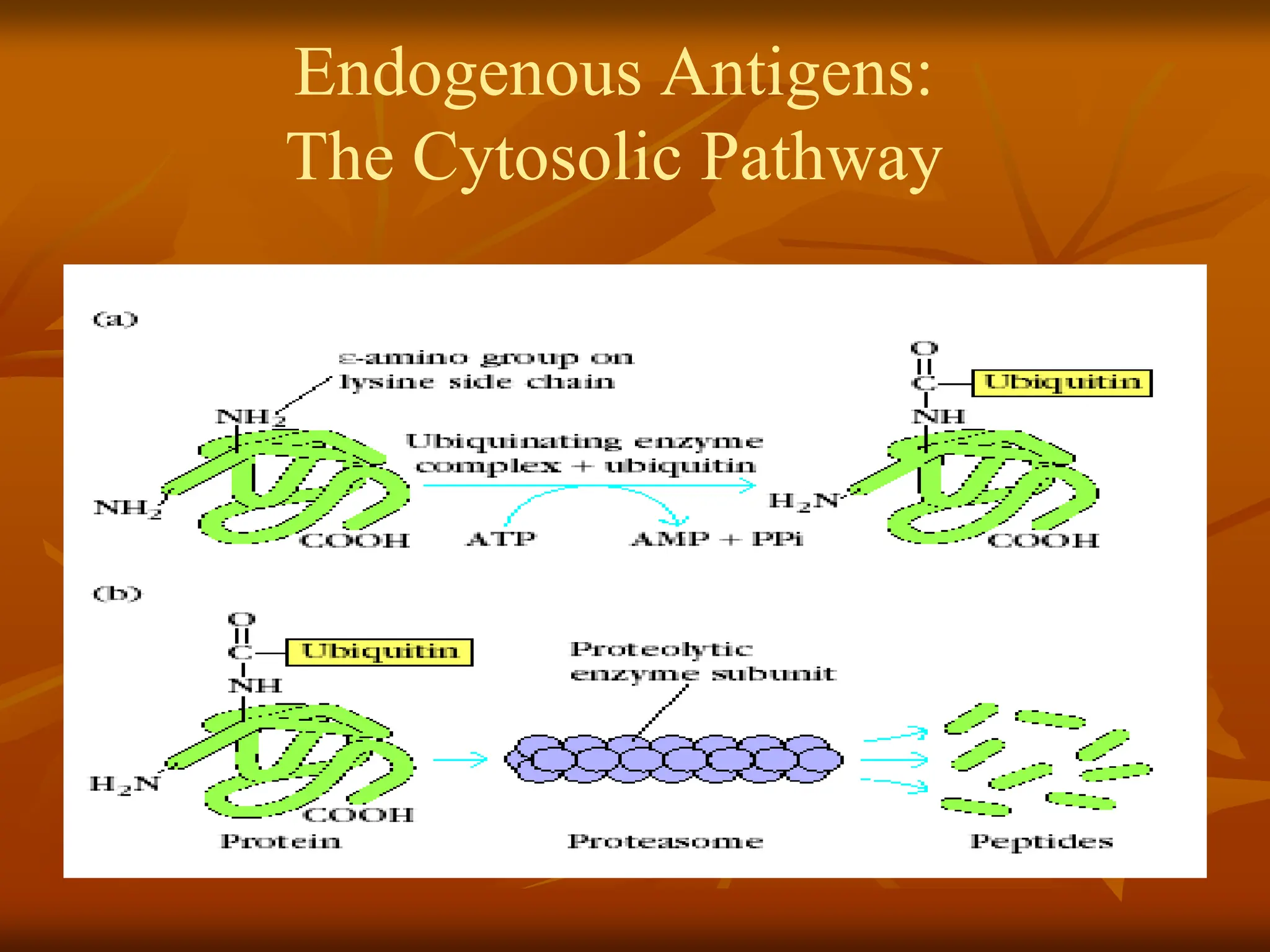
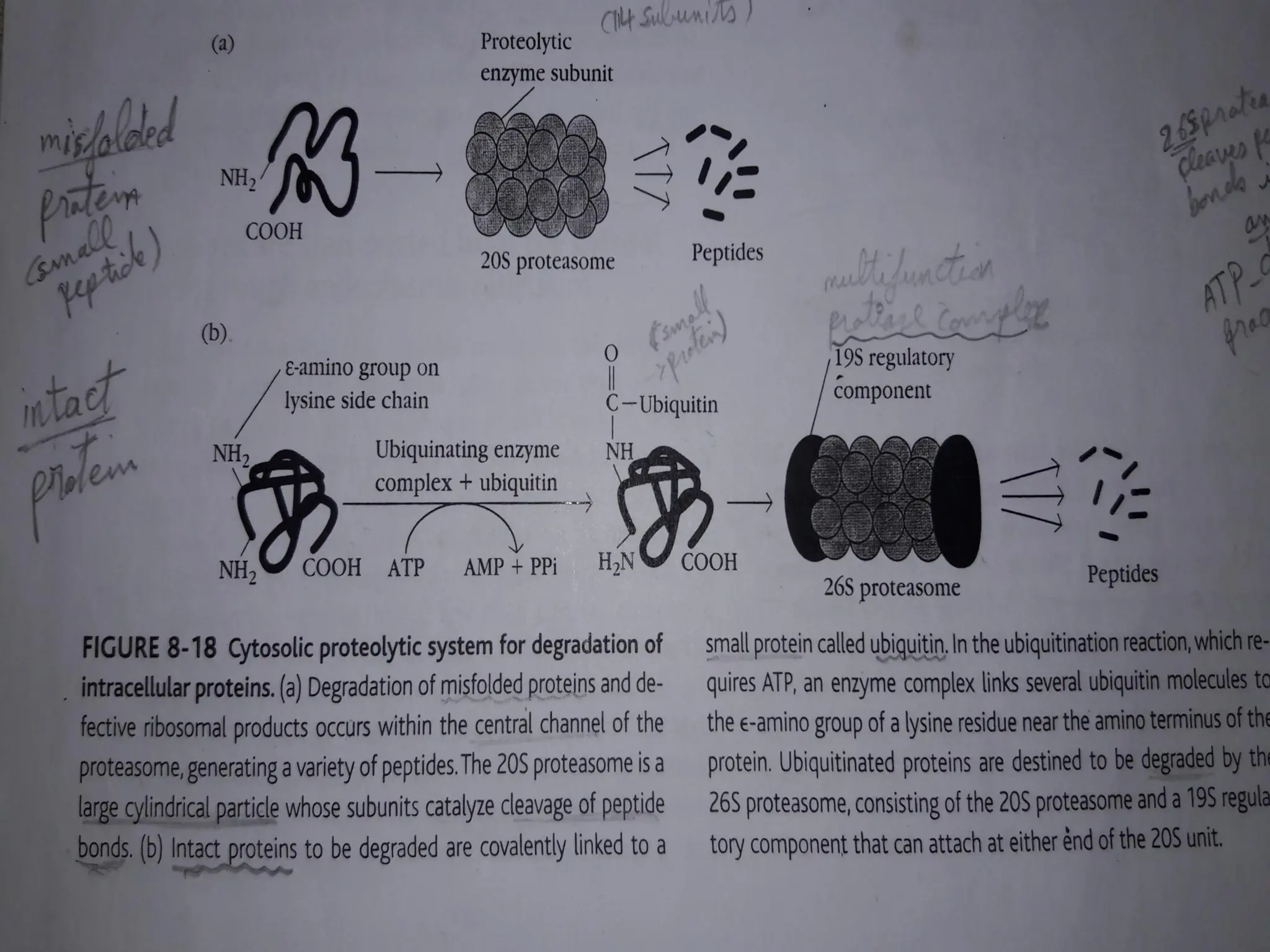
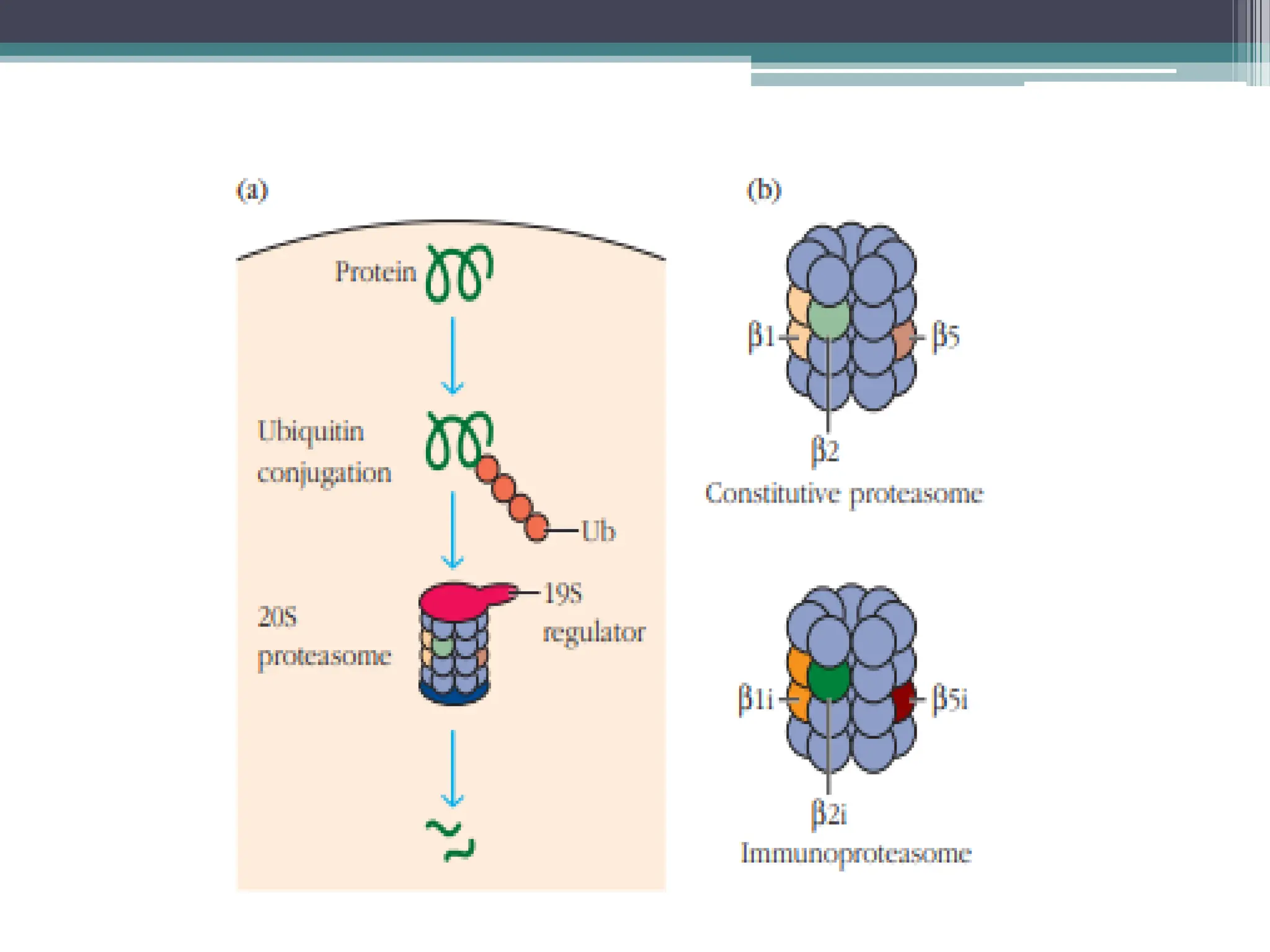
FIGURE - Cytosolicproteolytic system for degradation
of intracellular proteins. (a) Endogenous proteins may
be targeted for degradation by ubiquitin conjugation.
These proteins are degraded by the 26S proteasome
complex, which includes the 20S constitutive
proteasome and a 19S regulator. (b) In activated APCs,
several proteins in the constitutive proteasome
1,
2, and
are replaced by proteins encoded by the
LMP genes and specific to the immunoproteasome
(
1i,
2i, and
5i). This immunoproteasome has
increased proteolytic efficiency for creating peptides
that can assemble with MHC class I molecules.
19.
•
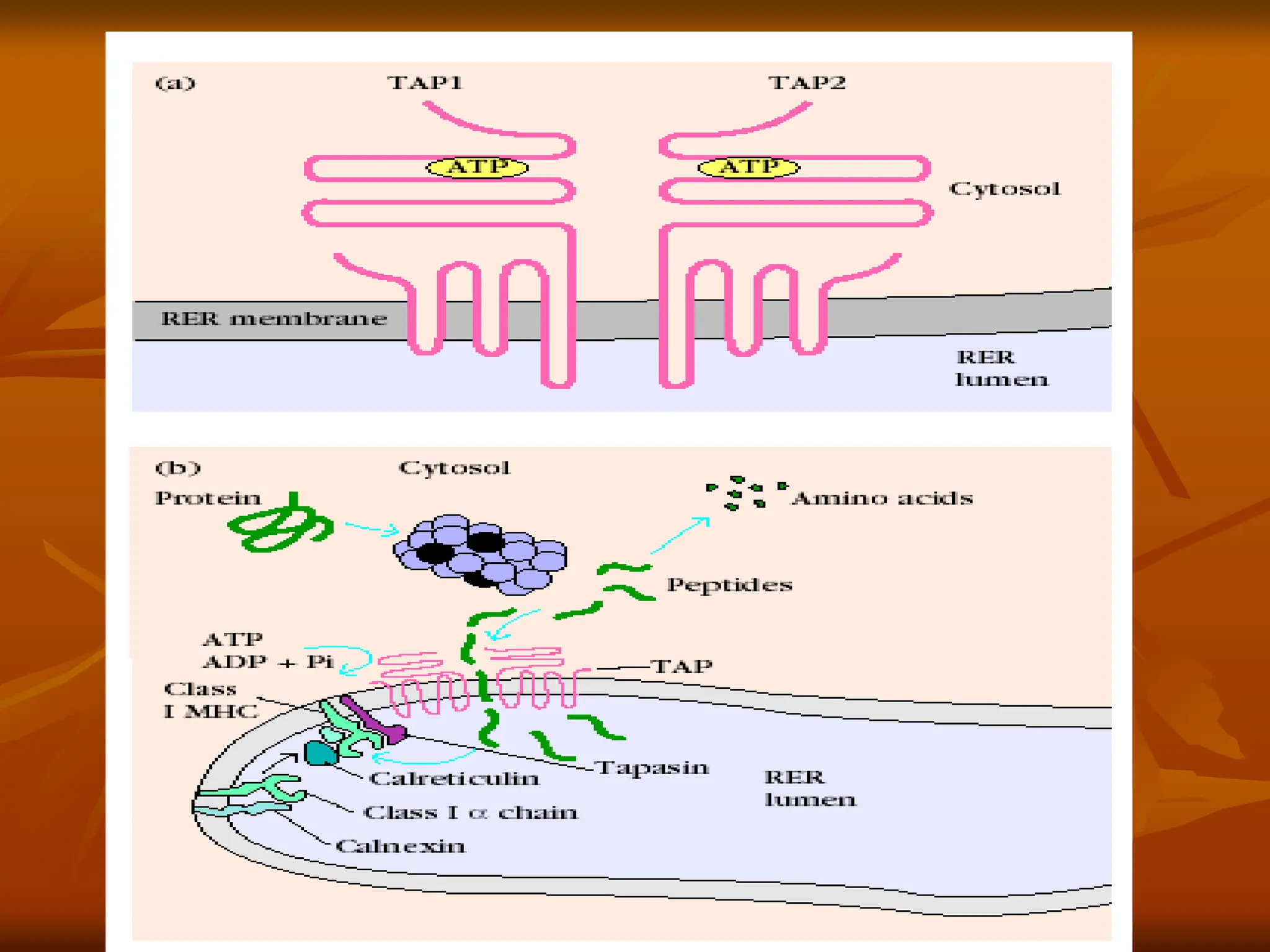
FIGURE - TAP(transporter associated with antigen
processing). (a) Schematic diagram of TAP, a
heterodimer anchored in the membrane of the rough
endoplasmic reticulum (RER). The two chains are
encoded by TAP1 and TAP2. The cytosolic domain in
each TAP subunit contains an ATP-binding site, and
peptide transport depends on the hydrolysis of ATP. (b)
In the cytosol, association of (colored spheres) with a
proteasome changes its catalytic specifi city to favor
production of peptides that bind to class I MHC
molecules. These peptides are translocated by TAP into
the RER lumen, where, in a process mediated by several
other proteins, they will associate with class I MHC
molecules.
22.
•
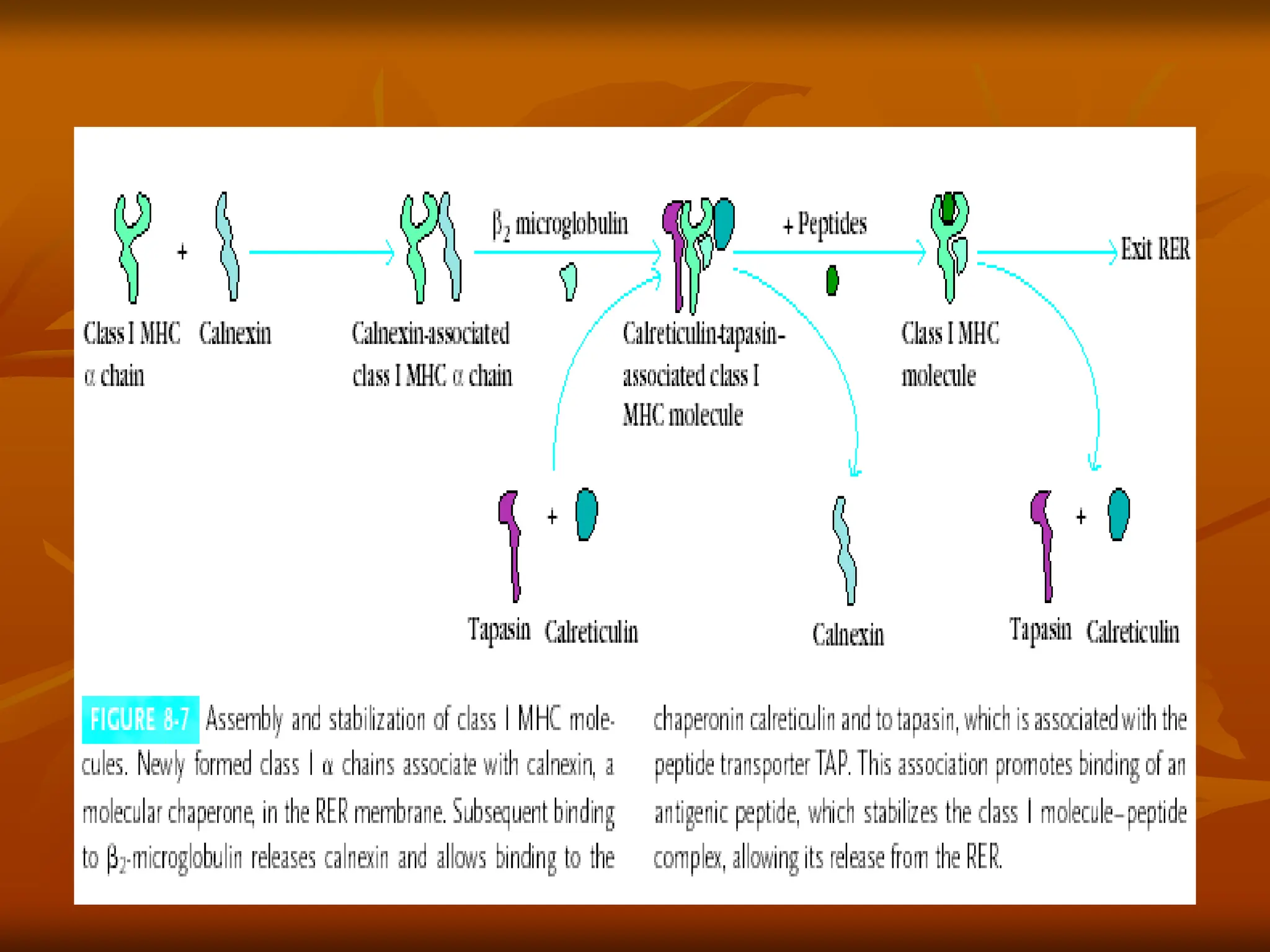
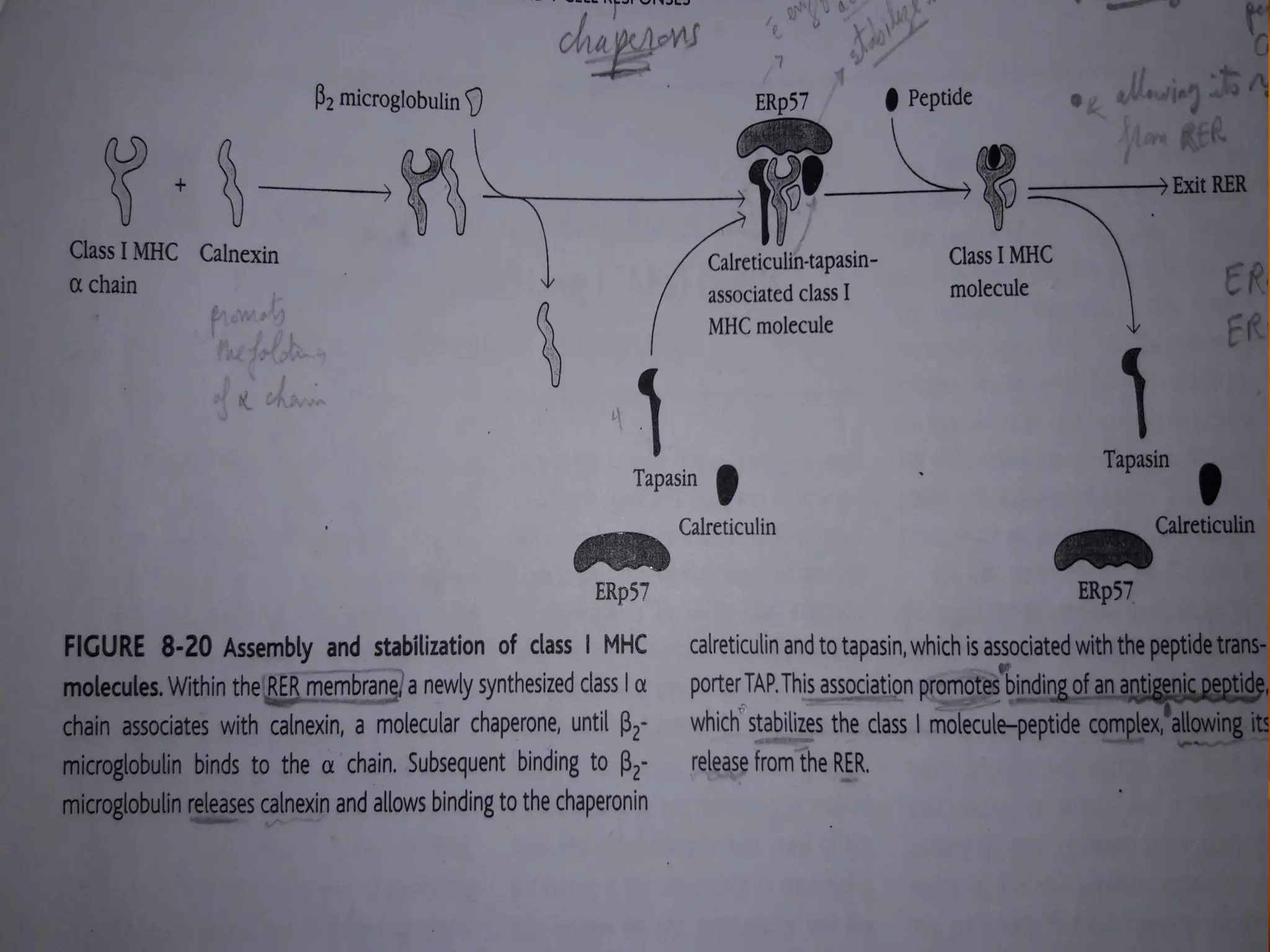
FIGURE - Assemblyand stabilization of class I MHC
•
molecules. Within the rough endoplasmic reticulum (RER) a
newly synthesized class I chain associates with Calnexin, a
molecular chaperone and ERp57 until
•
chain. The binding of
2-microglobulin binds to
•
2-microglobulin releases calnexin and
•
allows binding to calreticulin and to tapasin, which is associated
•
with the peptide transporter TAP. This association promotes
binding of an antigenic peptide. Antigens in the ER can be further
processed via exopeptidases such as ERAP1, producing fragments
ideally suited for binding to class I. Peptide association stabilizes the
class I molecule-peptide complex, allowing it to be transported from
the RER to the plasma membran.
•
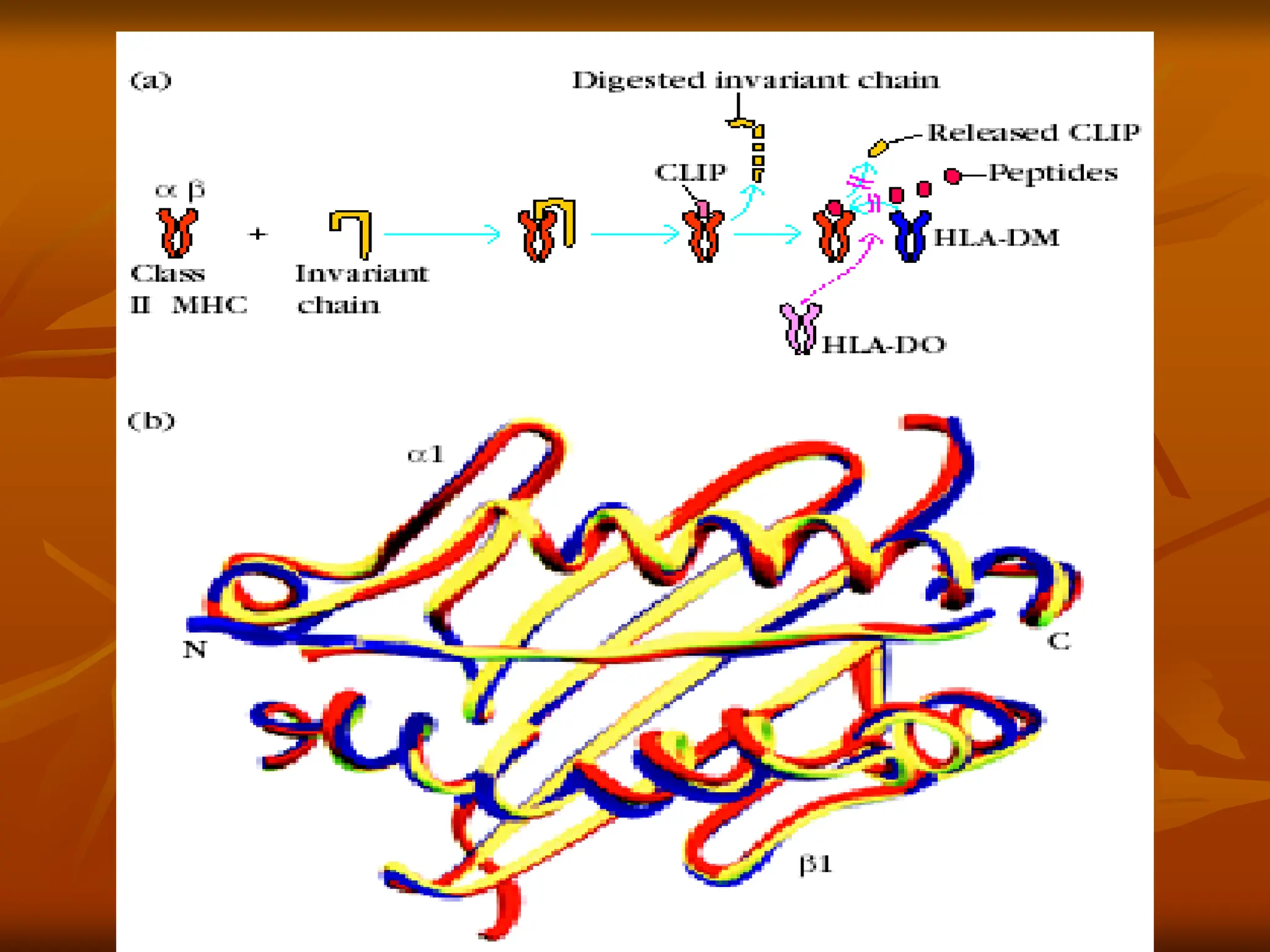
FIGURE - Assemblyof class II MHC molecules. (a) Within
•
the rough endoplasmic reticulum, a newly synthesized class II MHC
•
molecule binds an invariant chain. The bound invariant chain
prevents premature binding of peptides to the class II molecule and
•
helps to direct the complex to endocytic compartments containing
•
peptides derived from exogenous antigens. Digestion of the
invariant chain leaves CLIP, a small fragment remaining in the
binding groove of the class II MHC molecule. HLA-DM, a
nonclassical MHC class II molecule present within the MIIC
compartment, mediates exchange of antigenic peptides for CLIP.
The nonclassical class II molecule HLA-DO may act as a negative
regulator of class II antigen processing by binding to HLA-DM and
inhibiting its role in the dissociation of CLIP
sociation of CLIP
from class II molecules.
•
(b)Comparison of three-dimensional structures showing the binding
groove of HLA class II molecules containing different antigenic
peptides or CLIP.
28.
•
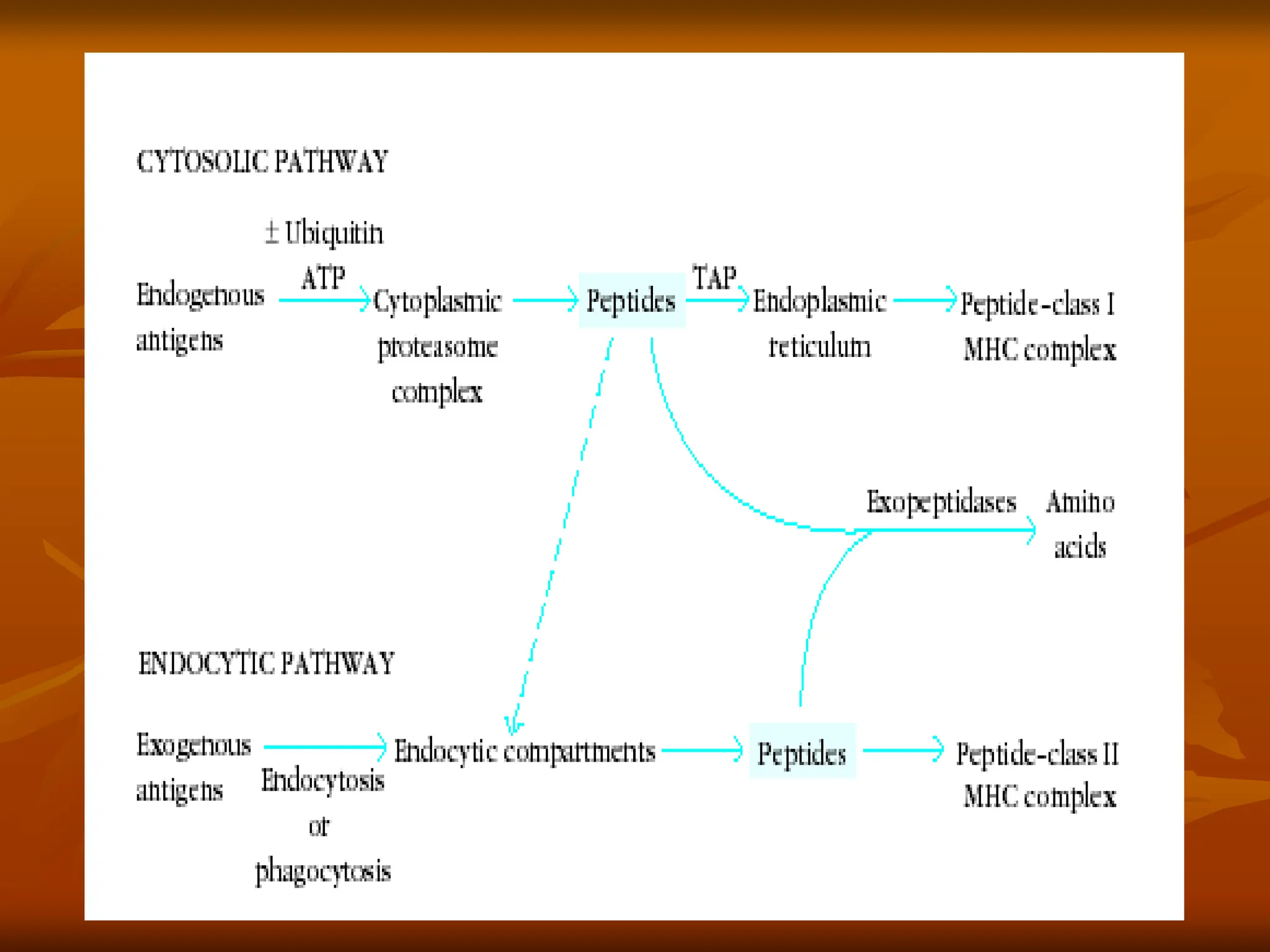
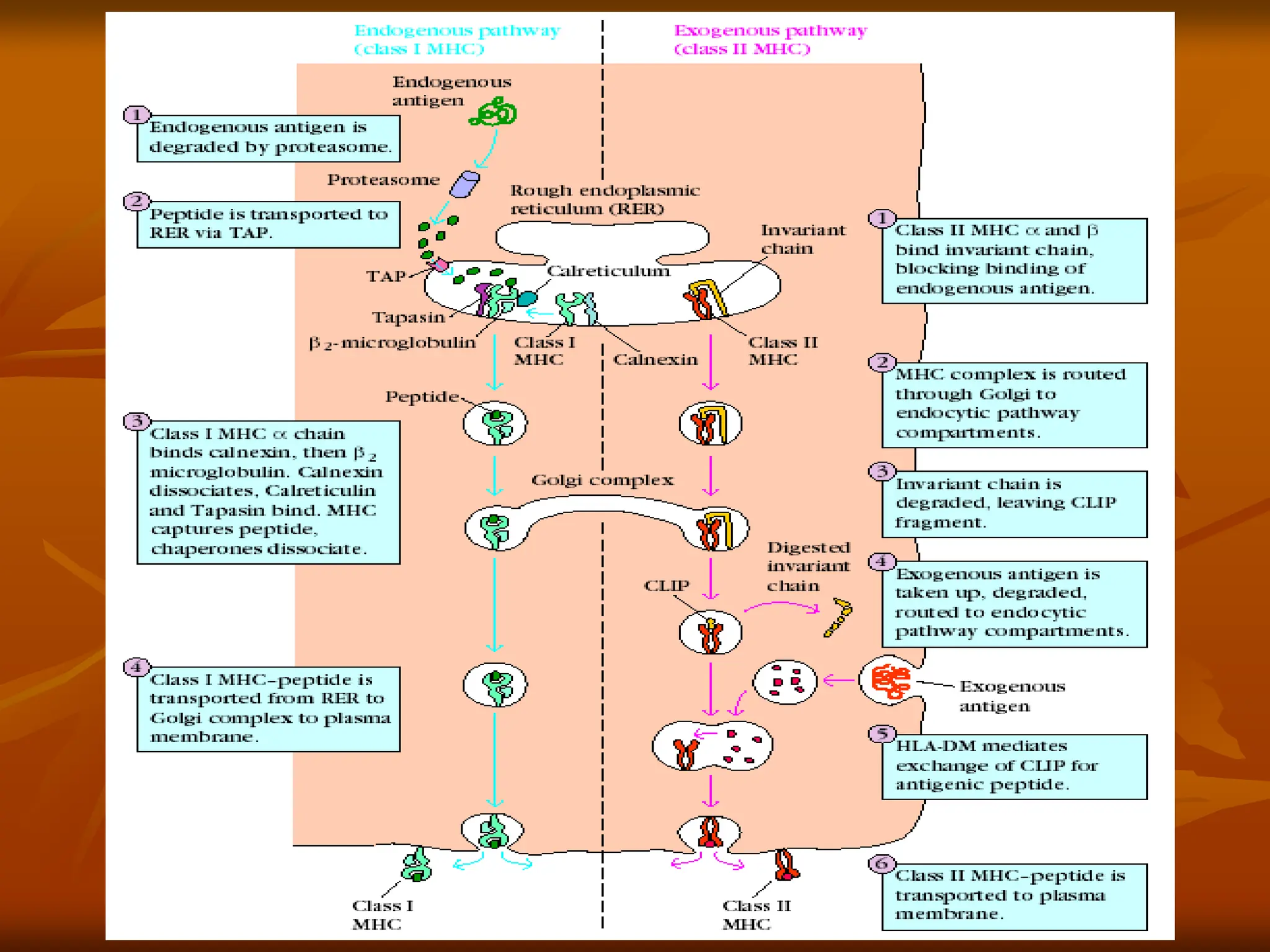
FIGURE - Overviewof endogenous and exogenous
pathways for processing antigen. In the
endogenous pathway (left), antigens are degraded by the
proteasome, converting proteins into smaller peptides.
In the exogenous pathway (right), extracellular antigens
are engulfed into endocytic compartments where they
are degraded by acidic pH-dependent endosomal and
lysosomal enzymes. The antigenic peptides from
proteasome cleavage and those from endocytic
compartments associate with class I or class II MHC
molecules respectively, and the MHC-peptide complexes
are then transported to the cell membrane. It should be
noted that the ultimate fate of most peptides in the cell is
neither of these pathways but rather to be degraded
completely into amino acids.
•
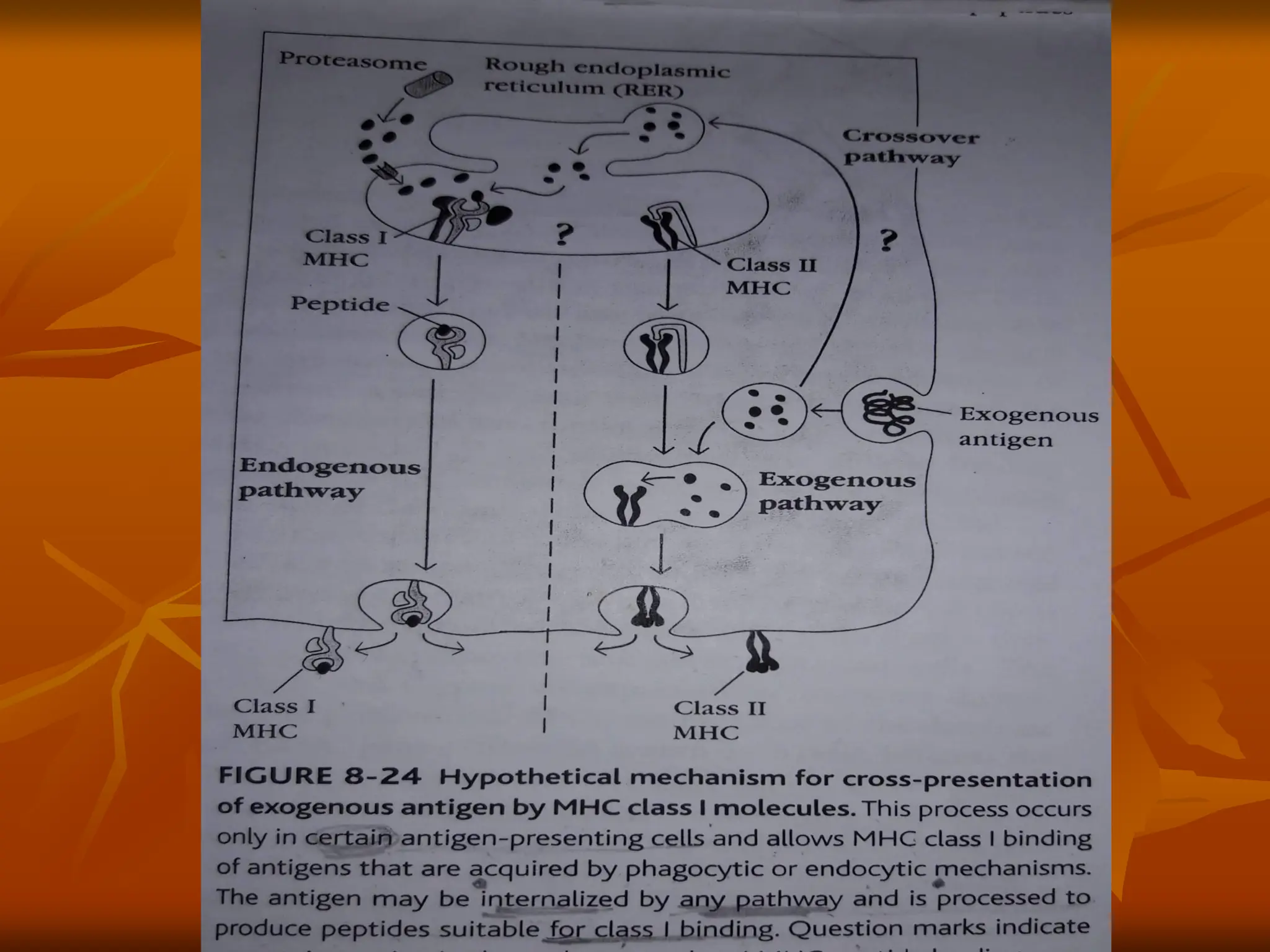
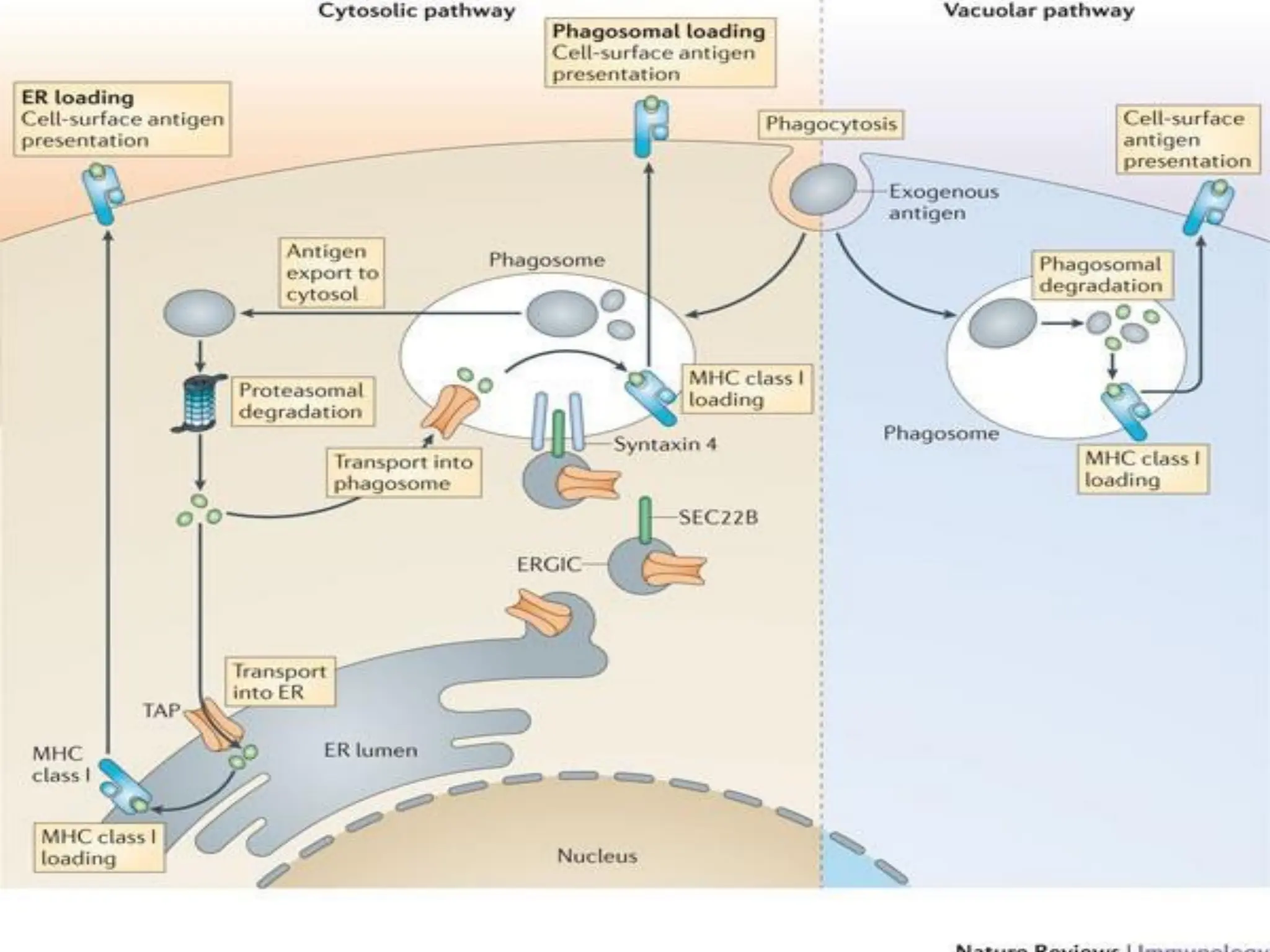
Figure 1 |Intracellular pathways for cross-presentation in
dendritic cells. After phagocytosis, exogenous antigens
can be exported into the cytosol, where they are
processed by the proteasome. The processed antigens
can then be loaded on MHC class I molecules in the
endoplasmic reticulum (ER) (the cytosolic pathway with
ER loading) or re-imported into the phagosome to be
loaded on MHC class I molecules (the cytosolic pathway
with phagosomal loading).
•
Alternatively, exogenous antigens can be degraded into
peptides in the phagosome, where they are then loaded
on MHC class I molecules (the vacuolar pathway).





















![ANTIGEN PROCESSING PRESENTATION AND RECOGNITION - Copy [Autosaved].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/antigenprocessingpresentationandrecognition-copyautosaved-220815200136-89a4c3c9-thumbnail.jpg?width=640&height=640&fit=bounds)








































![[DSC Europe 25] Dobrica Cosic - From Electrons to Innovation: How Granular Da...](https://cdn.slidesharecdn.com/ss_thumbnails/h4qk69zereaumbceubgr-dobrica-cosic-from-electrons-to-innovation-how-granular-data-and-analytics-are--251218085301-b982fb14-thumbnail.jpg?width=640&height=640&fit=bounds)
![[DSC Europe 25] Marko Djordjevic - AI can help Agriculture.pptx](https://cdn.slidesharecdn.com/ss_thumbnails/c0huq0ztiubmgccem2hc-marko-djordjevic-ai-can-help-agriculture-251218125253-7606f036-thumbnail.jpg?width=640&height=640&fit=bounds)
![[DSC Europe 25] Tatevik Maytesyan - How to actually use AI in marketing: gett...](https://cdn.slidesharecdn.com/ss_thumbnails/tjo626lsqdgfntbgl2mw-4-251216103155-e36cd239-thumbnail.jpg?width=640&height=640&fit=bounds)




![[DSC Europe 25] Hans Kleinsman - The Compliance Gearbox: How Tax Tech Mediate...](https://cdn.slidesharecdn.com/ss_thumbnails/dxdytie1toel0hr90bjs-2-251212103250-174fdbe7-thumbnail.jpg?width=640&height=640&fit=bounds)
![[DSC Europe 25] Maria Kokiasmenos - AI Governance US Perspective.pptx](https://cdn.slidesharecdn.com/ss_thumbnails/eszqnbzlsqa2vch6dmci-6-251215095918-6fcdf45f-thumbnail.jpg?width=640&height=640&fit=bounds)





![[DSC Europe 25] Debmalya Biswas - Agentification: the art of transforming man...](https://cdn.slidesharecdn.com/ss_thumbnails/r5azlggvtqiaiiusrqdr-4-251212103249-5a12c89b-thumbnail.jpg?width=640&height=640&fit=bounds)


