• An antigenis classically defined as a
molecular species/substance capable of
inducing immune response on introduction
into a suitable host and of reacting
specifically with products (antibody,
sensitized T-cells) manufactured as a
consequence of the immune response
• Exogenous Antigen
It is produced outside the cell & enters
the cell by endocytosis.
Presented by MHC class II molecules
Examples: Bacterial antigen, foreign
protiens.
3.
• Endogenous Antigen
Originateswithin the cell itself,
processed within cytoplasm of the cell
Presented by MHC Class I molecule
e.g. viral protein, tumor antigen
4.
• Antigen processingrefers to the ability
of antigen presenting cells (APCs) to
break down a protein antigen into
peptides and to associate those peptides
with MHC molecules
• Antigen presentation is the process of
displaying peptide antigens associated
with MHC molecules to a T cell
• Antigen presenting cells- Cells that
display peptides associated with class II
MHC molecules to CD4+ Th cells
5.
•Target Cells- Cellsthat display peptides
associated with class I MHC molecules to
CD8+ Tc cells
•Self MHC Restriction- T cells from any
individual recognize peptide antigens only
when these peptides are bound to &
displayed by MHC molecules of that
individual
7.
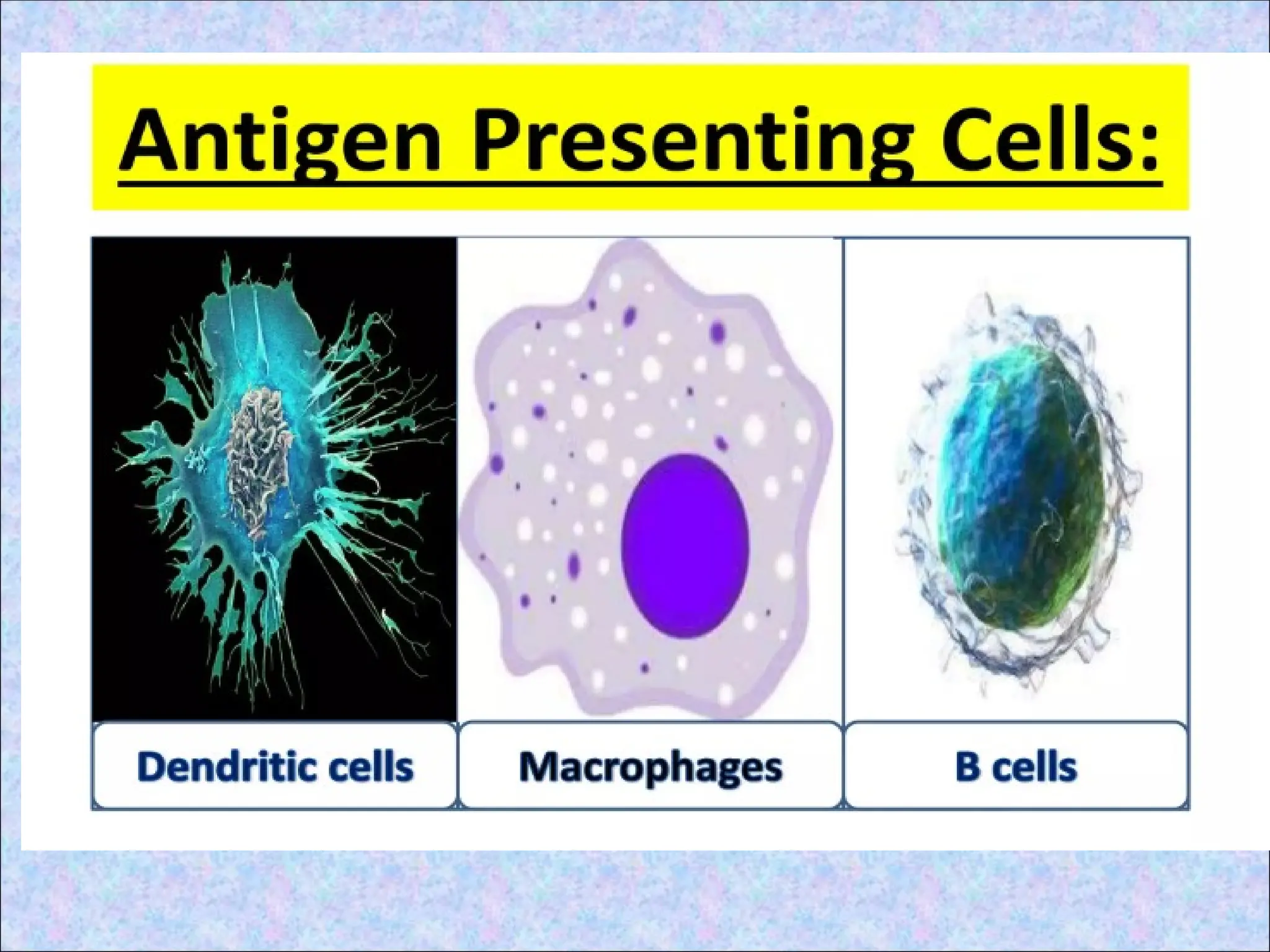
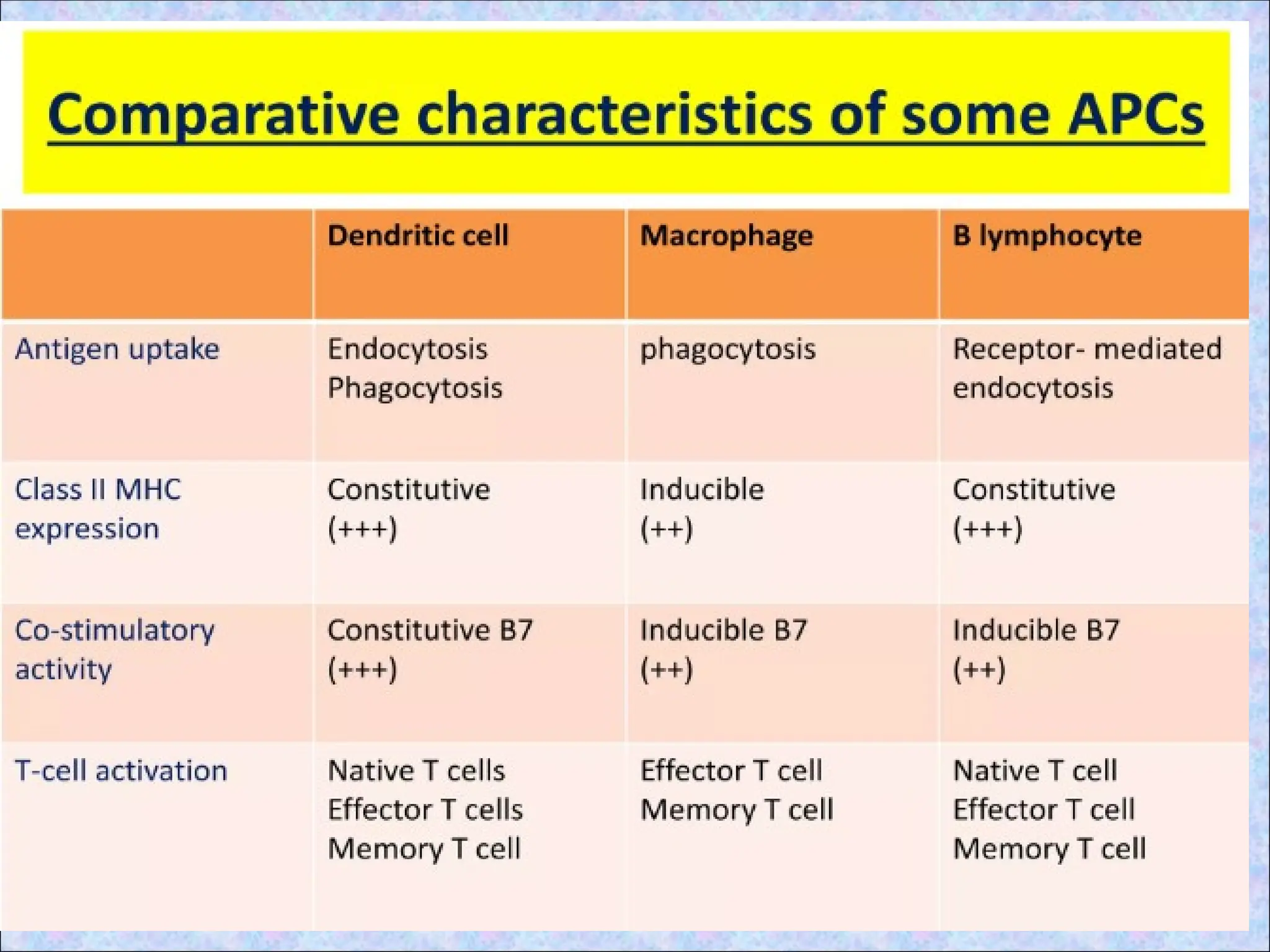
ANTIGEN PRESENTING
CELLS(APCs)
• Fewnucleated cells having MHC II
• Capture and display the antigens to T cells
• Provide accessory stimulus; costimulator
• Professional APCs Non-professional
• Dendritic cells constitutive
• B cells
• Macrophages Non-constitutive
11.
DENDRITIC CELLS:
• Haveno known function other than antigen
presentation to T cells
• 100 times more potent in this role than
macrophages and B cells
• Most effective APC for activation of
“naïve” T cells as express high levels of
both class II MHC molecules and members
of the co-stimulatory B7 family
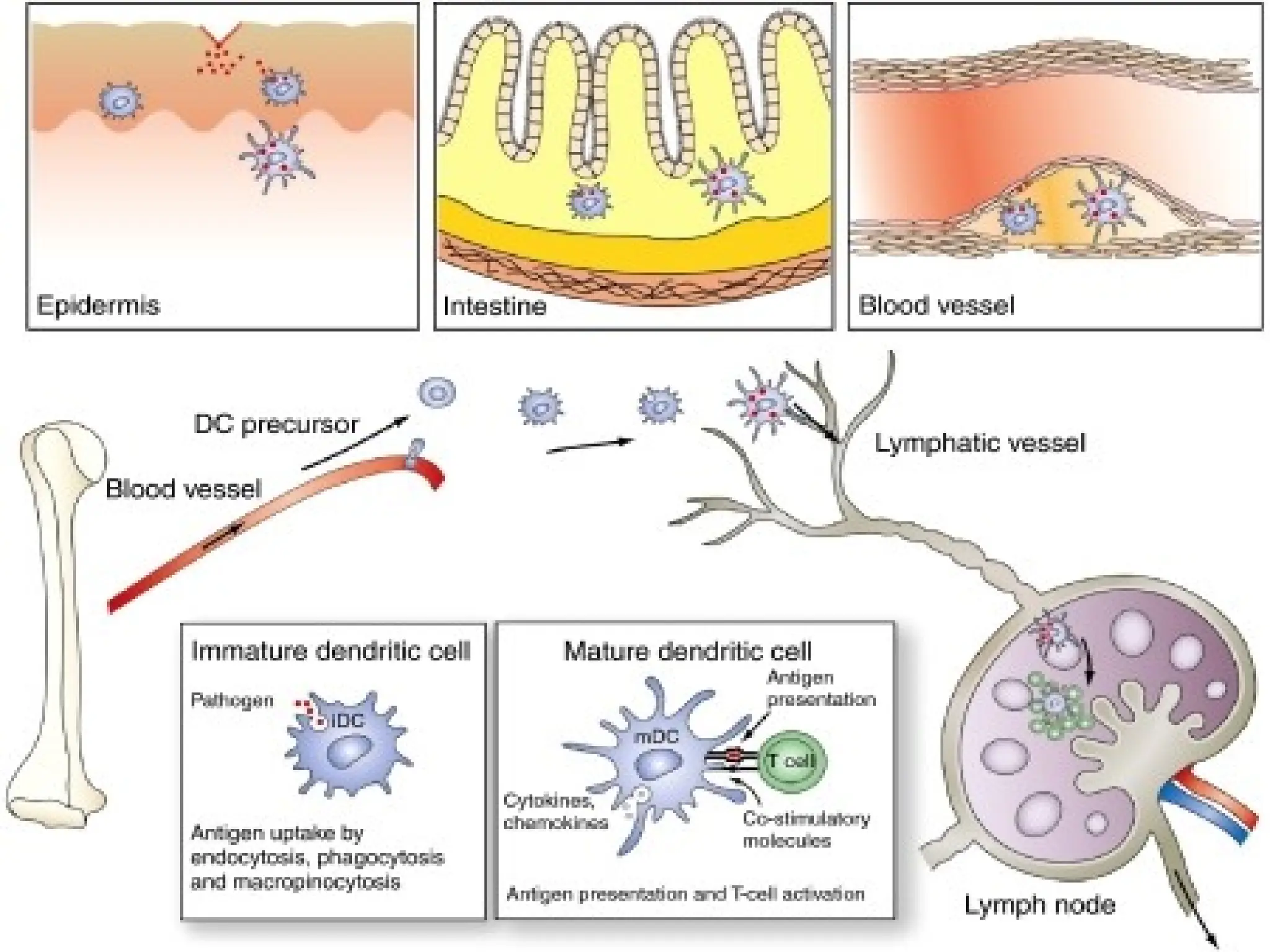
• Origin:
Produced from bone marrow stem cells
Immature dendritic cells then migrate
around the body & form lattice-like
networks in almost every tissue
12.
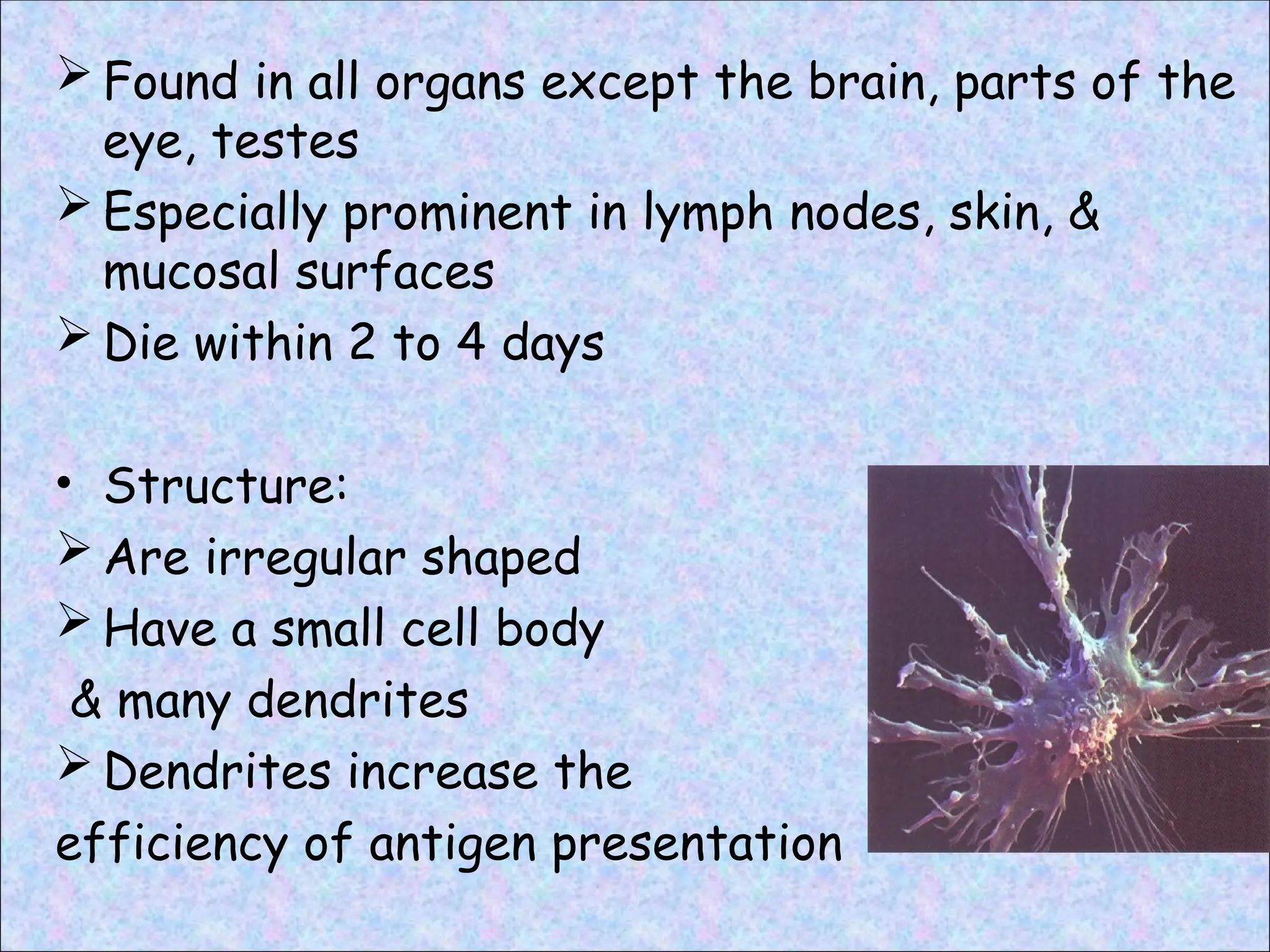
Found inall organs except the brain, parts of the
eye, testes
Especially prominent in lymph nodes, skin, &
mucosal surfaces
Die within 2 to 4 days
• Structure:
Are irregular shaped
Have a small cell body
& many dendrites
Dendrites increase the
efficiency of antigen presentation
MACROPHAGES
• Actively phagocytose
largeparticles.
• Play important role in
presenting antigens
derived from certain
microorganism such as
bacteria & parasites.
Not as effective as DCs
in activating “naïve” T
cells
• Effective in activating
memory T cells(effector
phase)
19.
• Expresses receptors
forFc portion of
antibodies and
complement protein C3b
which bind antigen
• IFN-γ secreted by T-
cells activate
macrophages to become
more effective killers
of microorganism.
20.
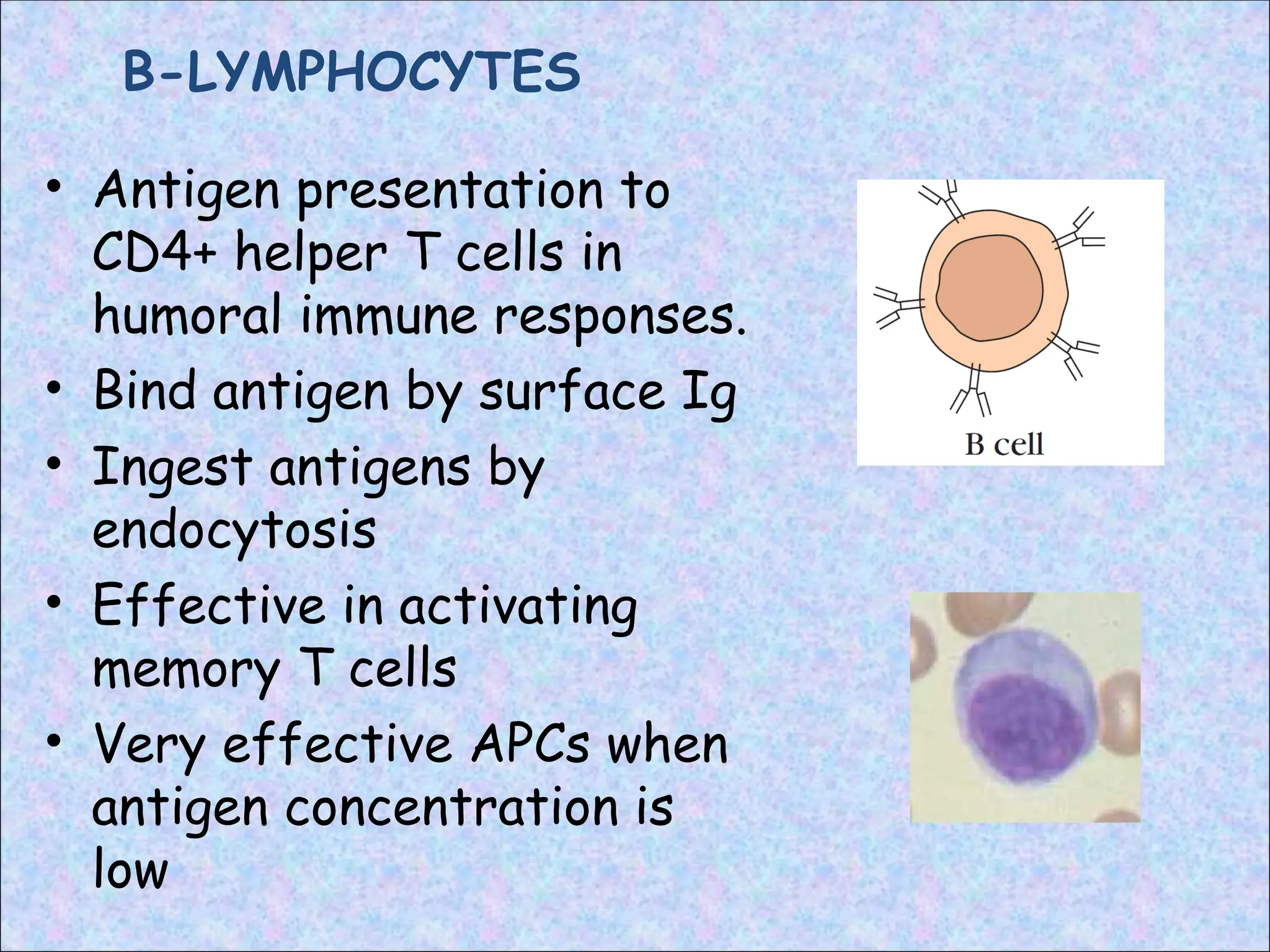
B-LYMPHOCYTES
• Antigen presentationto
CD4+ helper T cells in
humoral immune responses.
• Bind antigen by surface Ig
• Ingest antigens by
endocytosis
• Effective in activating
memory T cells
• Very effective APCs when
antigen concentration is
low
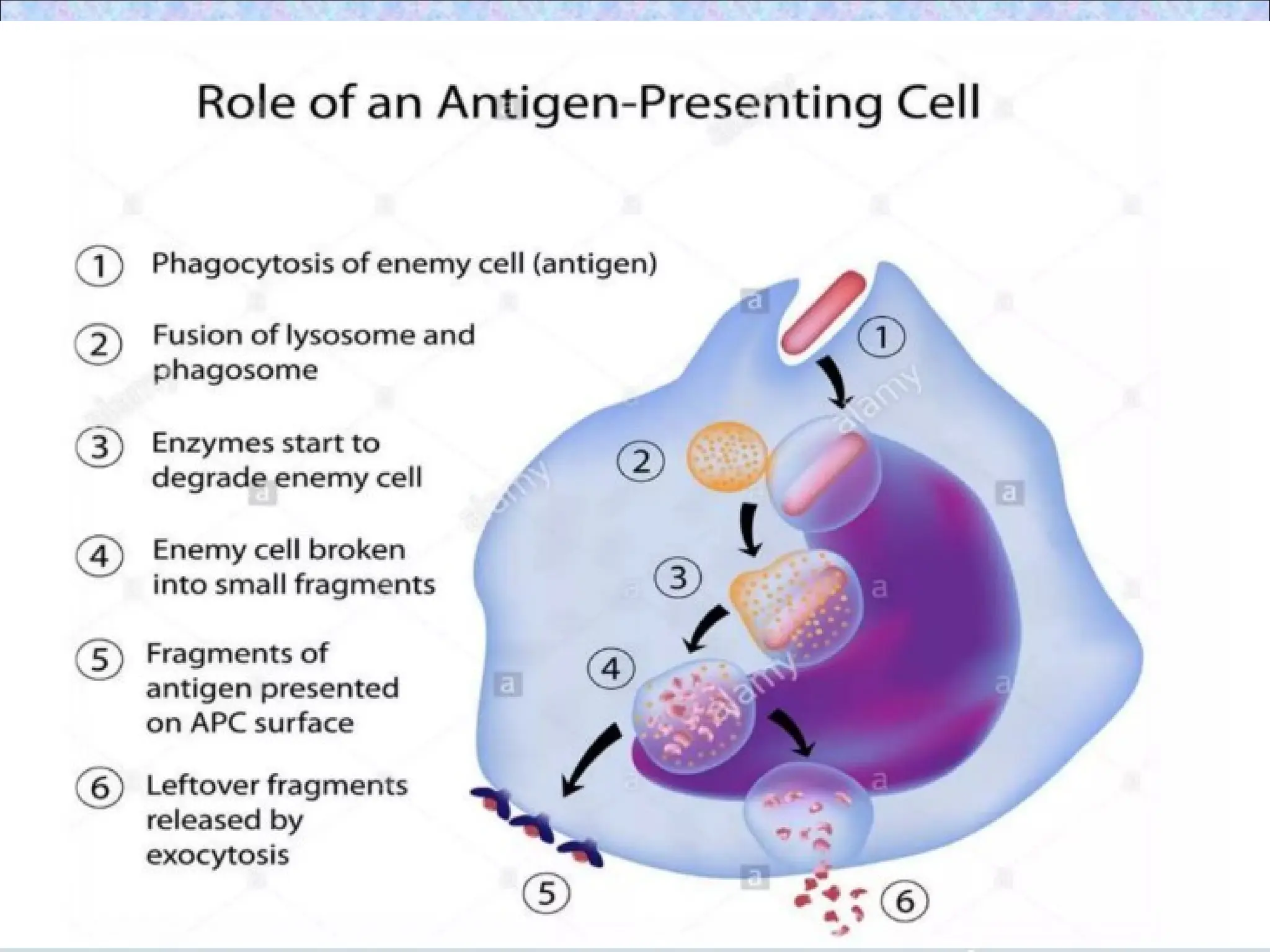
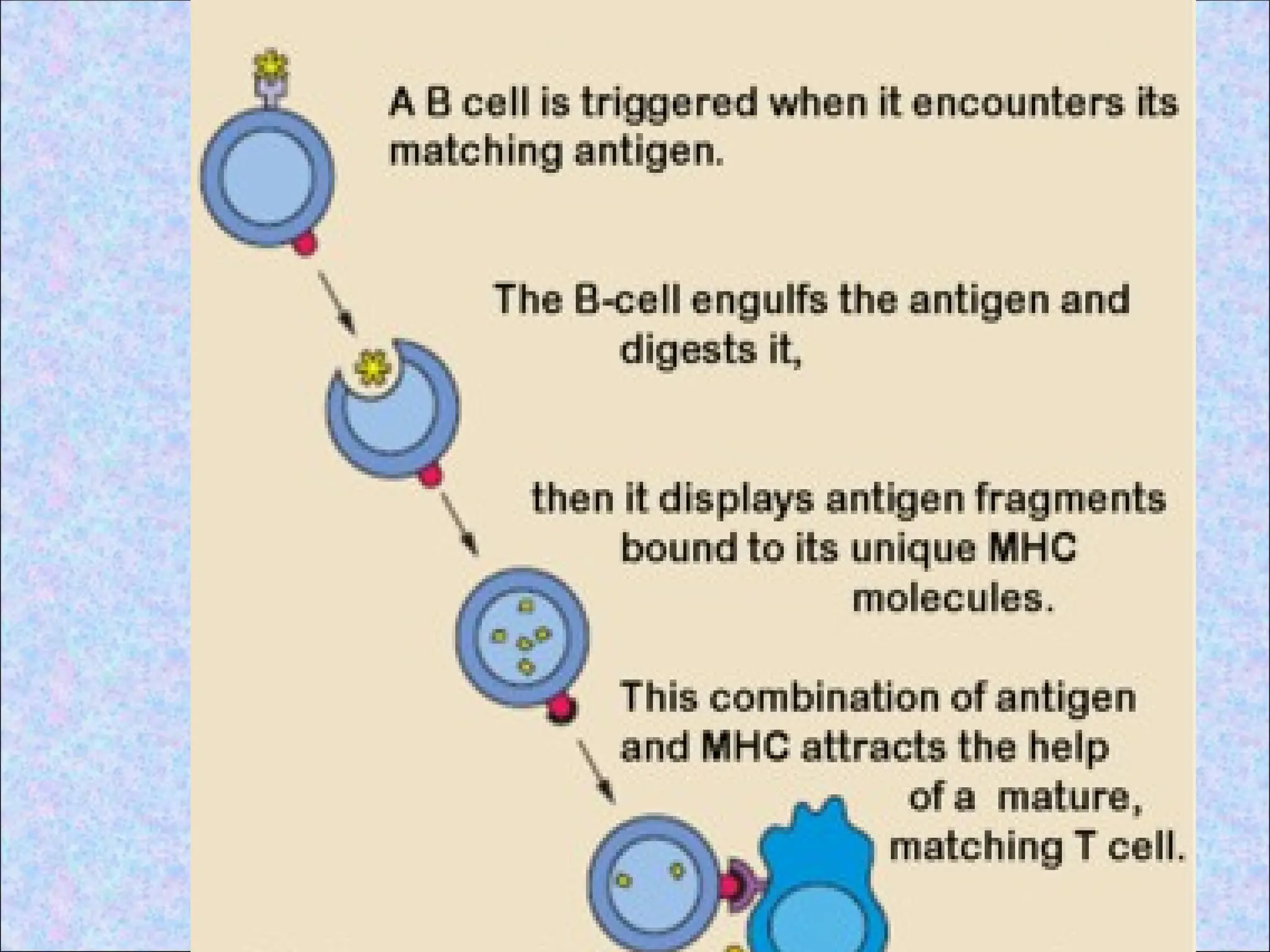
• Involves proteolyticdegradation of
internalized proteins in endocytic vesicles
& binding of peptides to class II MHC
molecules in these vesicles
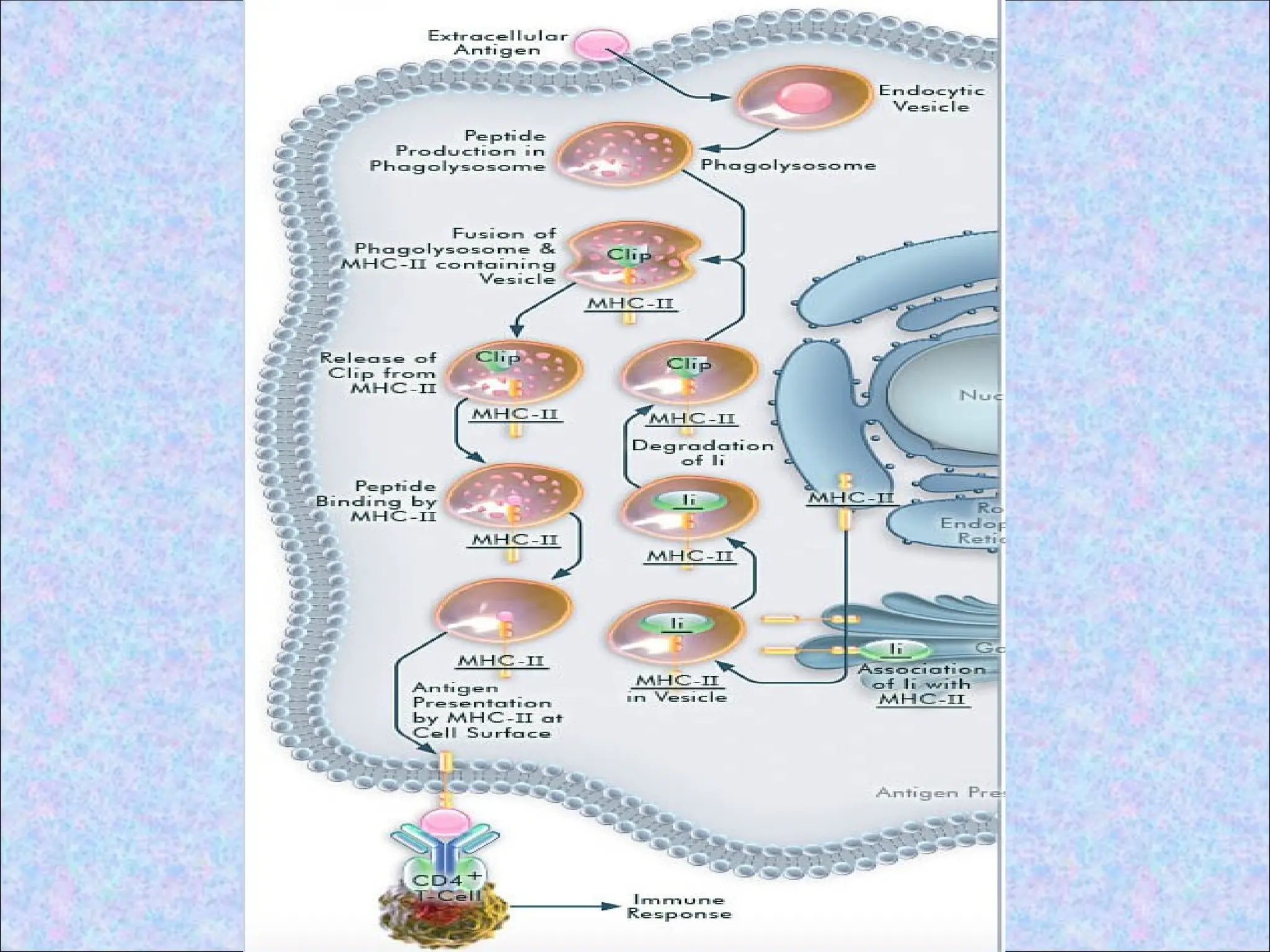
1.Uptake of extracellular proteins into
vesicular compartment of APCs
• Native antigen binds to an APC and
internalization of antigen occurs
• After internalization protein antigen
becomes localized in cellular membrane
bound vesicles (endosomes)
24.
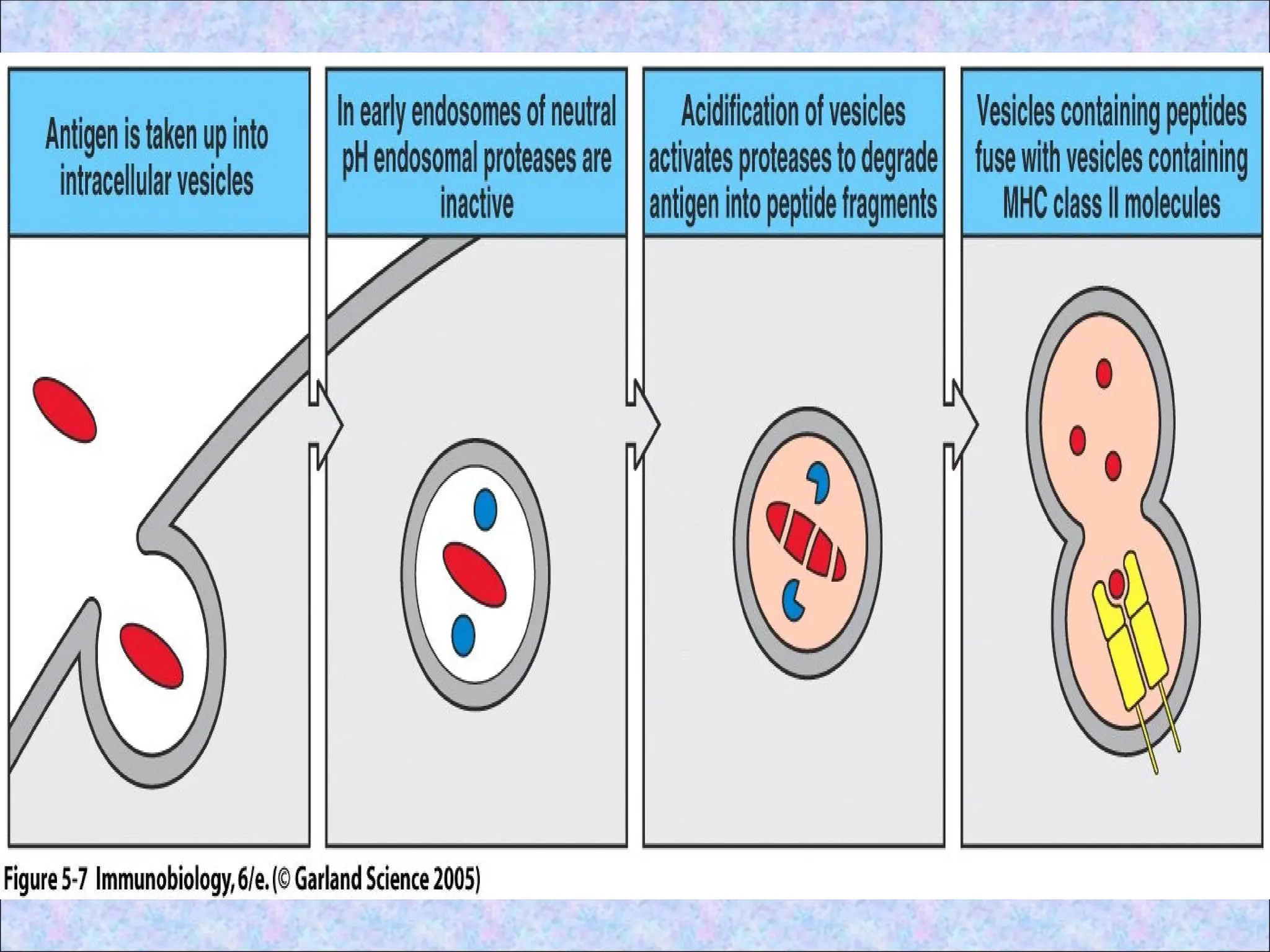
• Endosomes communicatewith lysosomes
2.Processing of internalised proteins in
endosomal and lysosomal vesicles
• Internalised proteins are degraded
enzymatically in late endosomes and
lysosomes to generate peptides which bind
to peptide clefts of class II MHC molecule
• Degradation of protein antigens is an
active process and mediated by proteases
• Most abundant proteases of endosomes
are cathepsins
26.
• Partially degradedproteins bind to the open-
ended clefts of class II MHC molecules and
are then trimmed enzymatically to final size
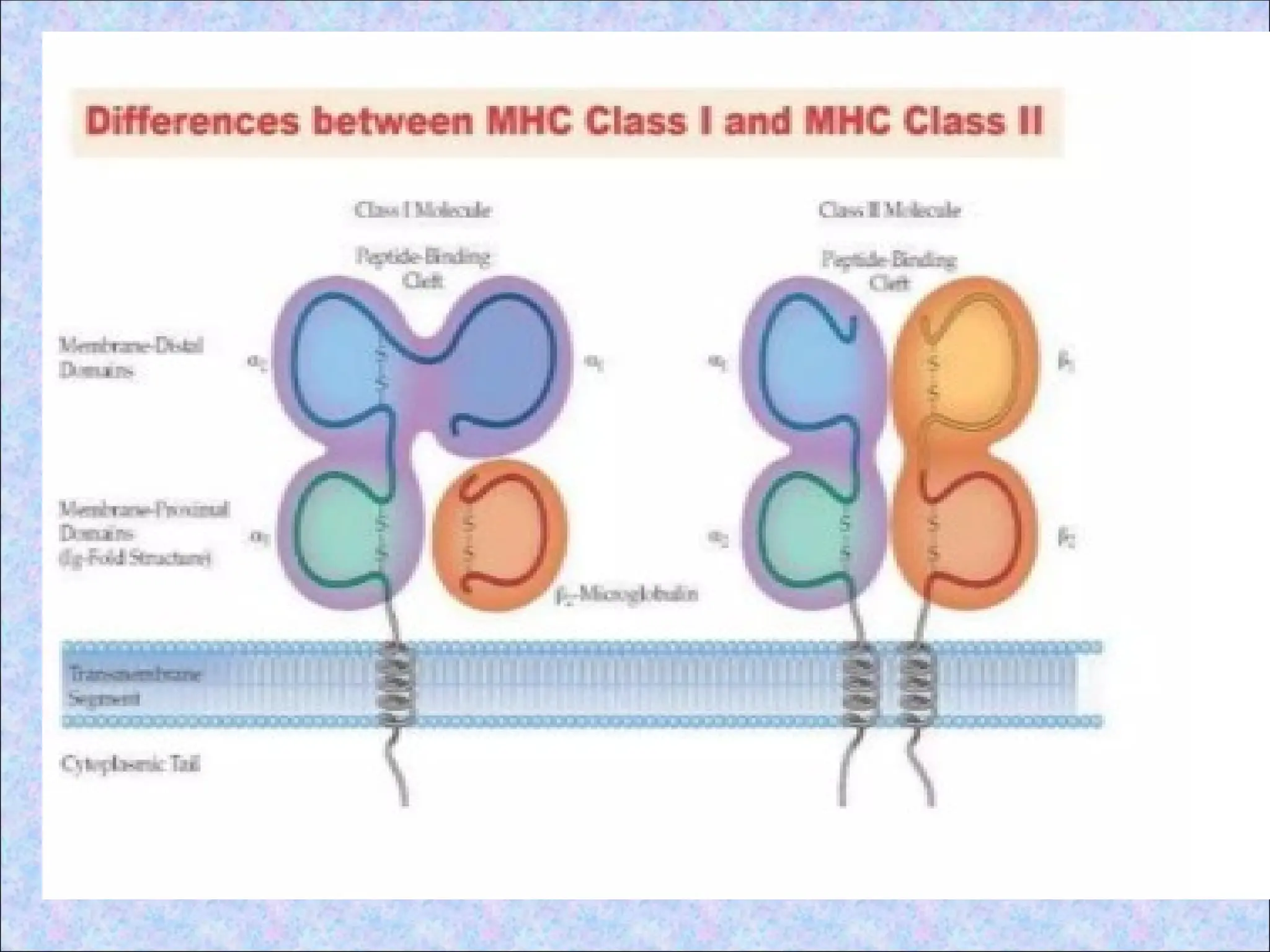
3.Biosynthesis and transport of class II MHC
molecules to endosomes
• Class II MHC molecules are synthesised in
the ER and transported to endosomes with
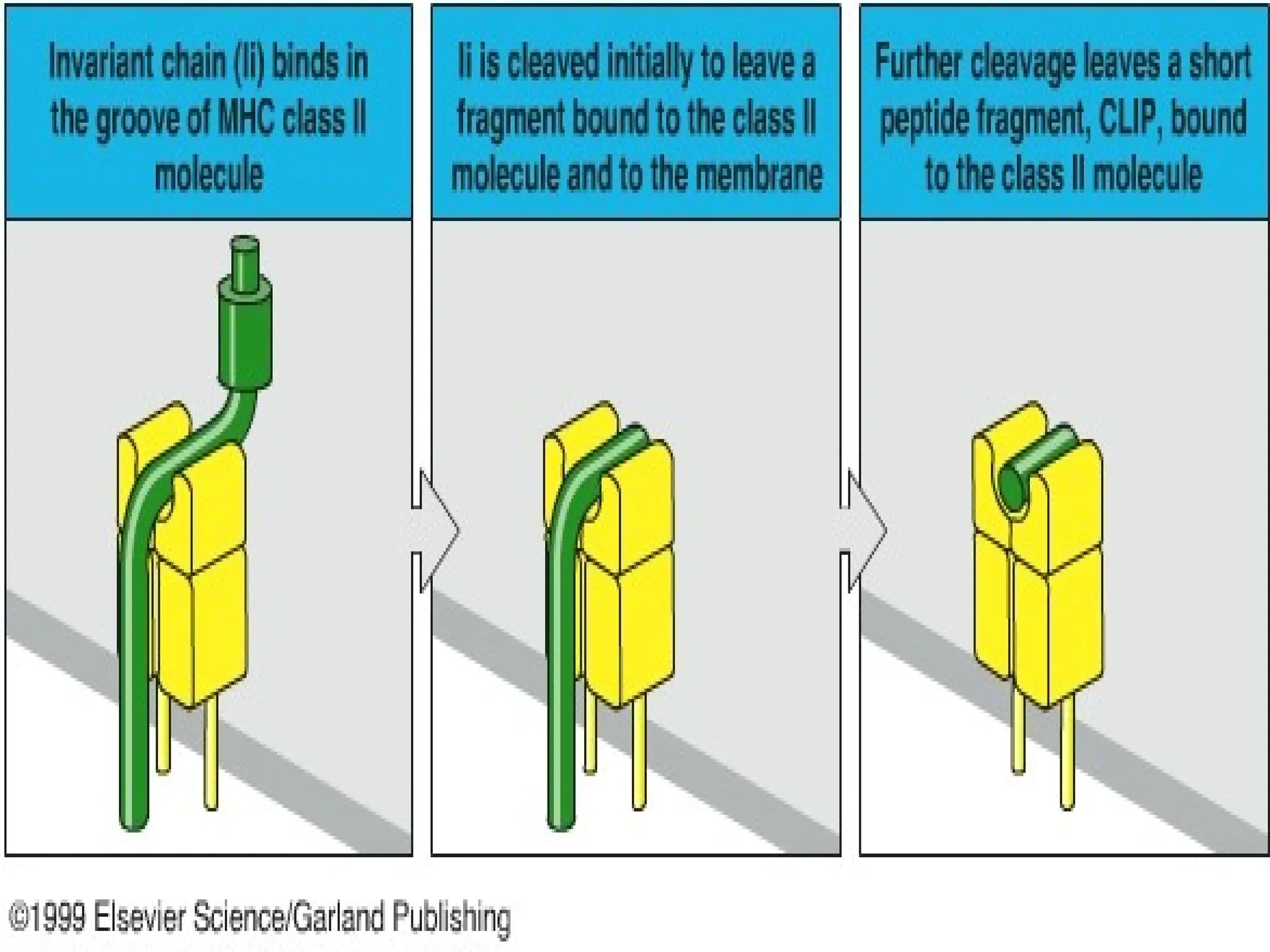
an associated protein (invariant chain ‘Ii’)
• Ii occupies the peptide binding clefts of the
newly synthesised class II molecules
• α and β chains are coordinately synthesised
and associate with each other in the ER
27.
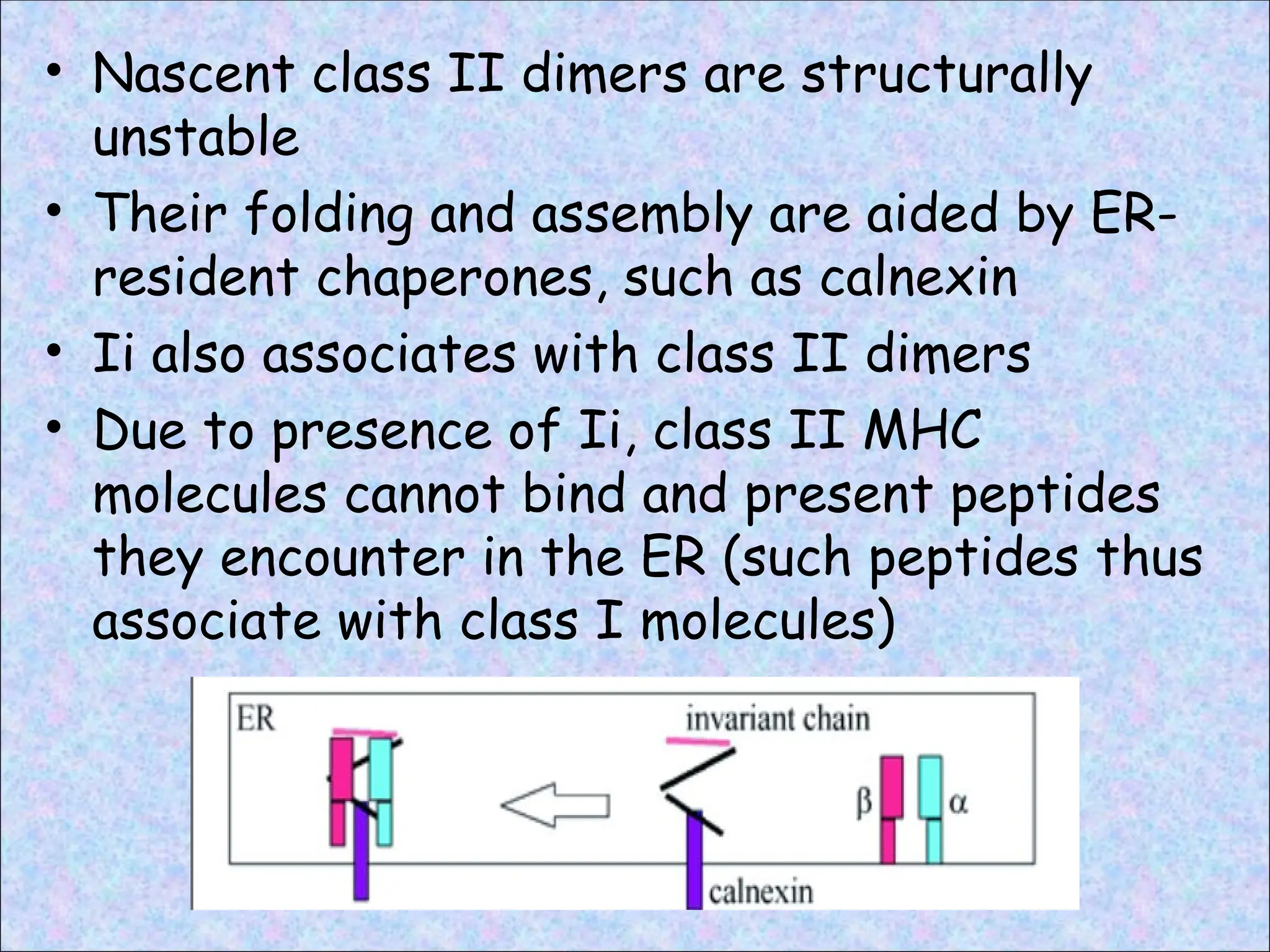
• Nascent classII dimers are structurally
unstable
• Their folding and assembly are aided by ER-
resident chaperones, such as calnexin
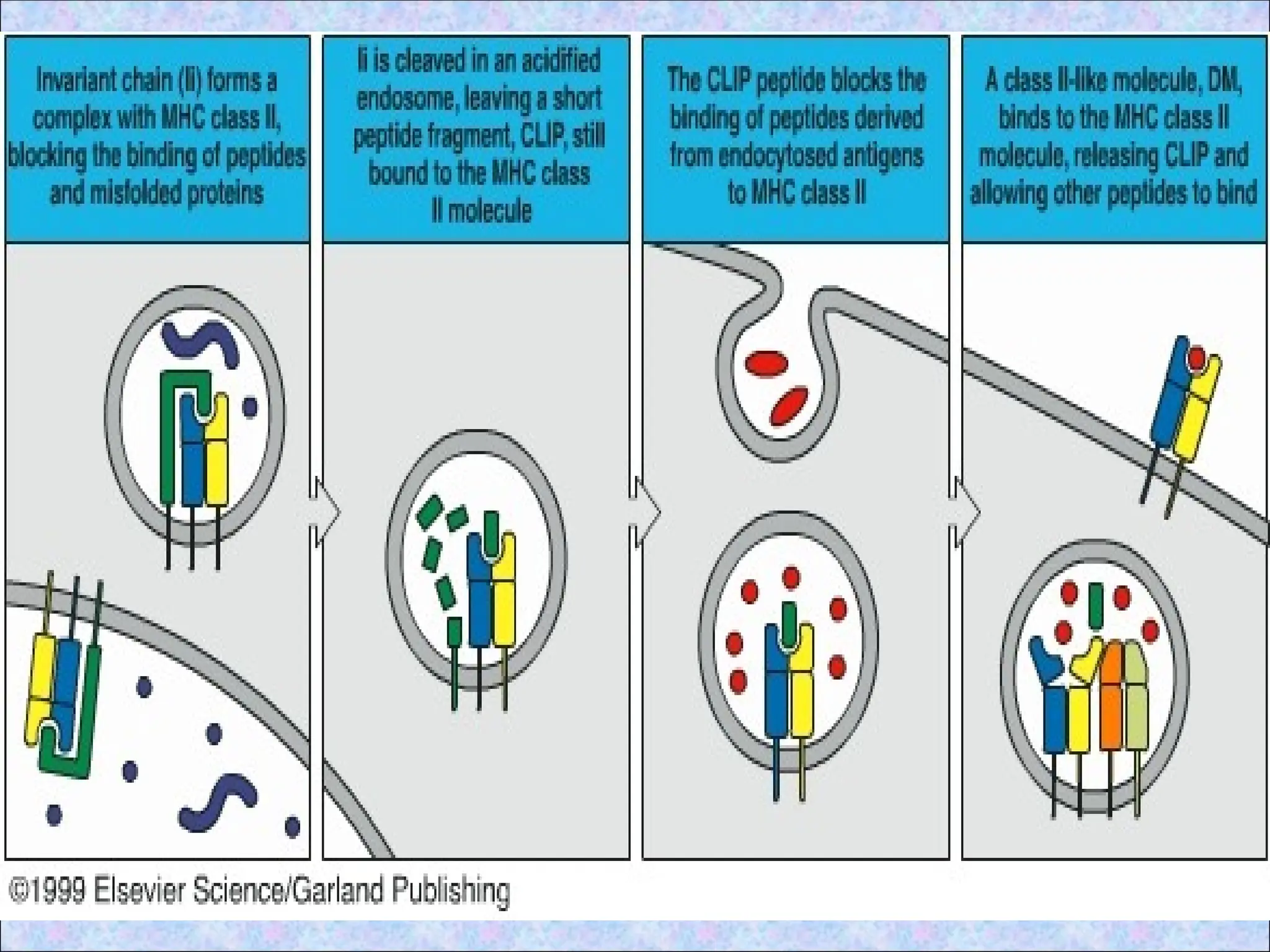
• Ii also associates with class II dimers
• Due to presence of Ii, class II MHC
molecules cannot bind and present peptides
they encounter in the ER (such peptides thus
associate with class I molecules)
28.
• Vesicles containingclass II molecules
emerge from Golgi complex and are
targeted to late endosomes and
lysosomes
• Exocytic vesicles fuse with endocytic
veiscles containing internalised and
processed antigens
• As a result, class II molecules enter the
vesicles containing peptides generated
by proteolysis of endocytosed proteins
29.
4. Association ofprocessed peptides with
class II MHC molecules in vesicles
• Ii dissociates from class II MHC
molecules by the combined action of
proteolytic enzymes
• Cathepsin S acts on Ii and degrades it
to form CLIP (Class II associated
invariant chain peptide)
• CLIP is a 24 amino acid long peptide
• Removal of CLIP is required before the
cleft becomes accessible to peptides
produced from extracellular proteins
31.
• Antigenic peptidesare then able to bind
to the available peptide-binding clefts
of the class II molecules
• HLA-DM aids in removal of CLIP
• Ends of the class II MHC peptide
binding cleft are open
• Bound peptides are trimmed by
proteolytic enzymes to the appropriate
size for T cell recognition
33.
5. Expression ofpeptide-class II complexes
on the APC surface
• Class II MHC molecules are stabilised by
the bound peptides
• The stable peptide-class II complexes
are delivered to the surface of APC,
where they are displayed for recognition
by CD4+ T cells
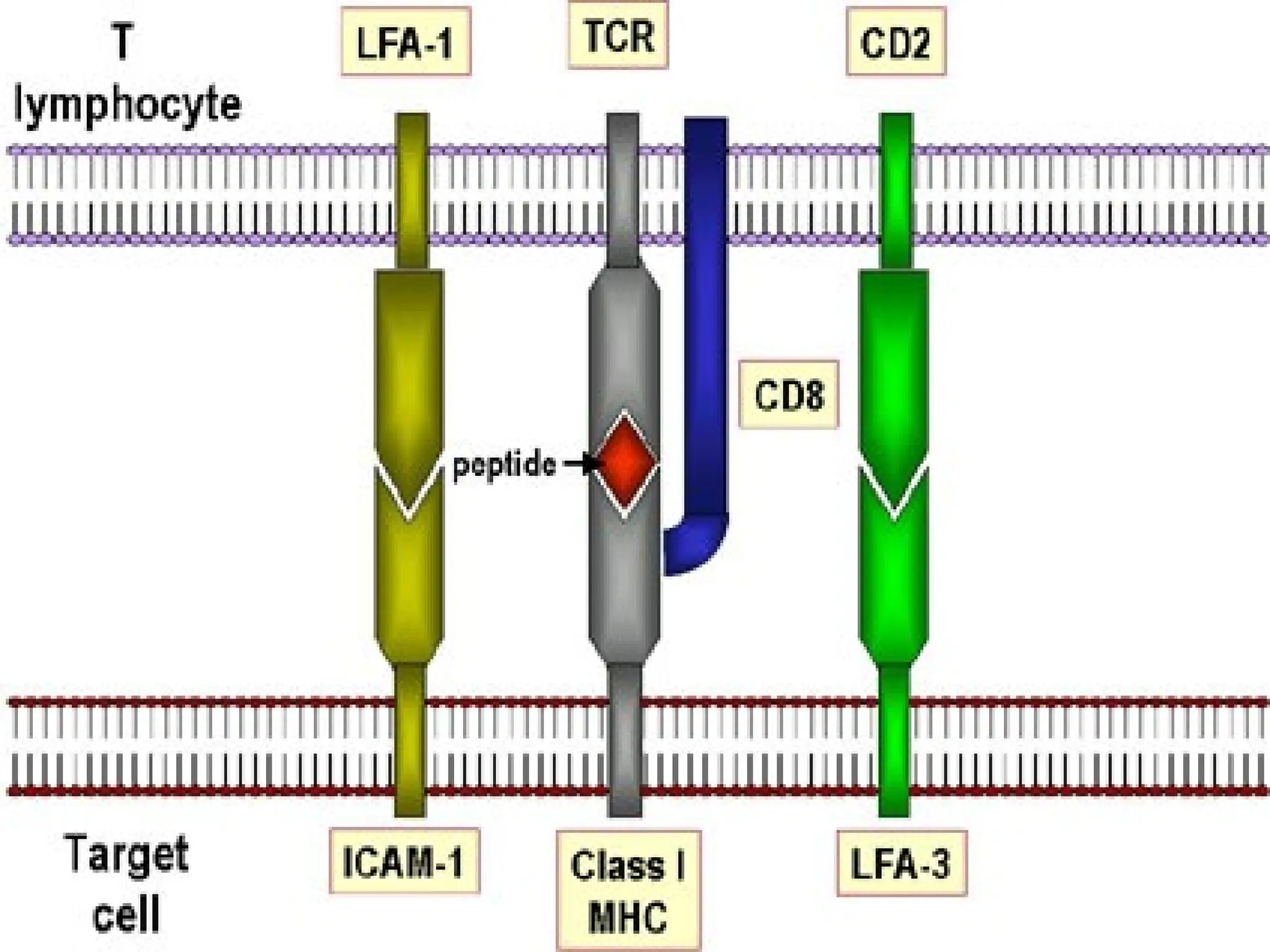
• CD4 co receptor plays an essential role in
recognition by CD4+ cells
35.
CO RECEPTOR: CD4
•Transmembrane glycoprotein
member of Ig superfamily
• Expressed as a monomer on the
surface of peripheral T cells
and thymocytes
• Also present on mononuclear
phagocytes & some dendritic
cells
• Has four extracellular Ig-like
domains, a hydrophobic
transmembrane region & a
highly basic cytoplasmic tail, 38
amino acids long
36.
• The twoN-terminal Ig-like domains of
the CD4 protein bind to the β2 domain
of the class II MHC molecule
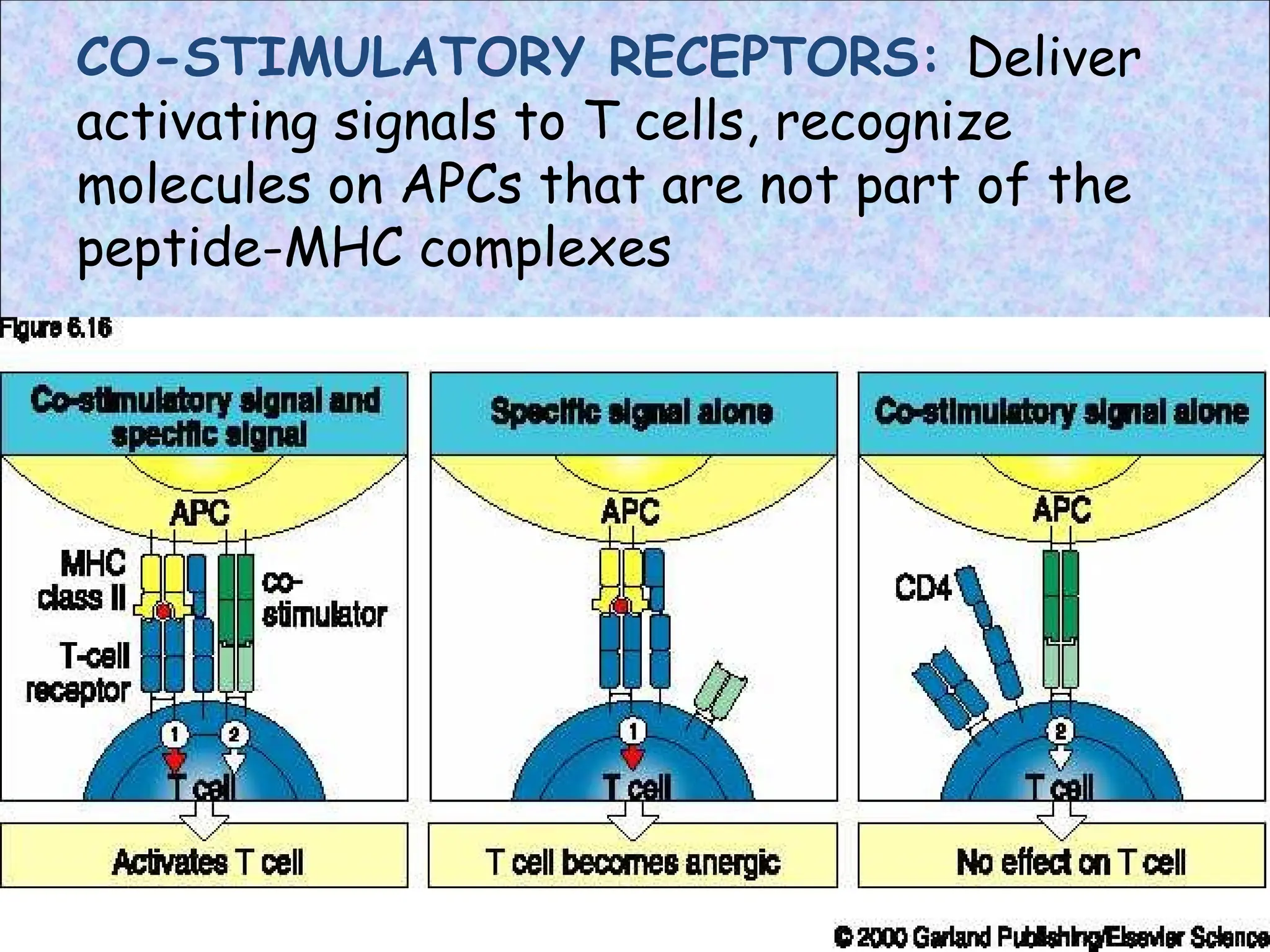
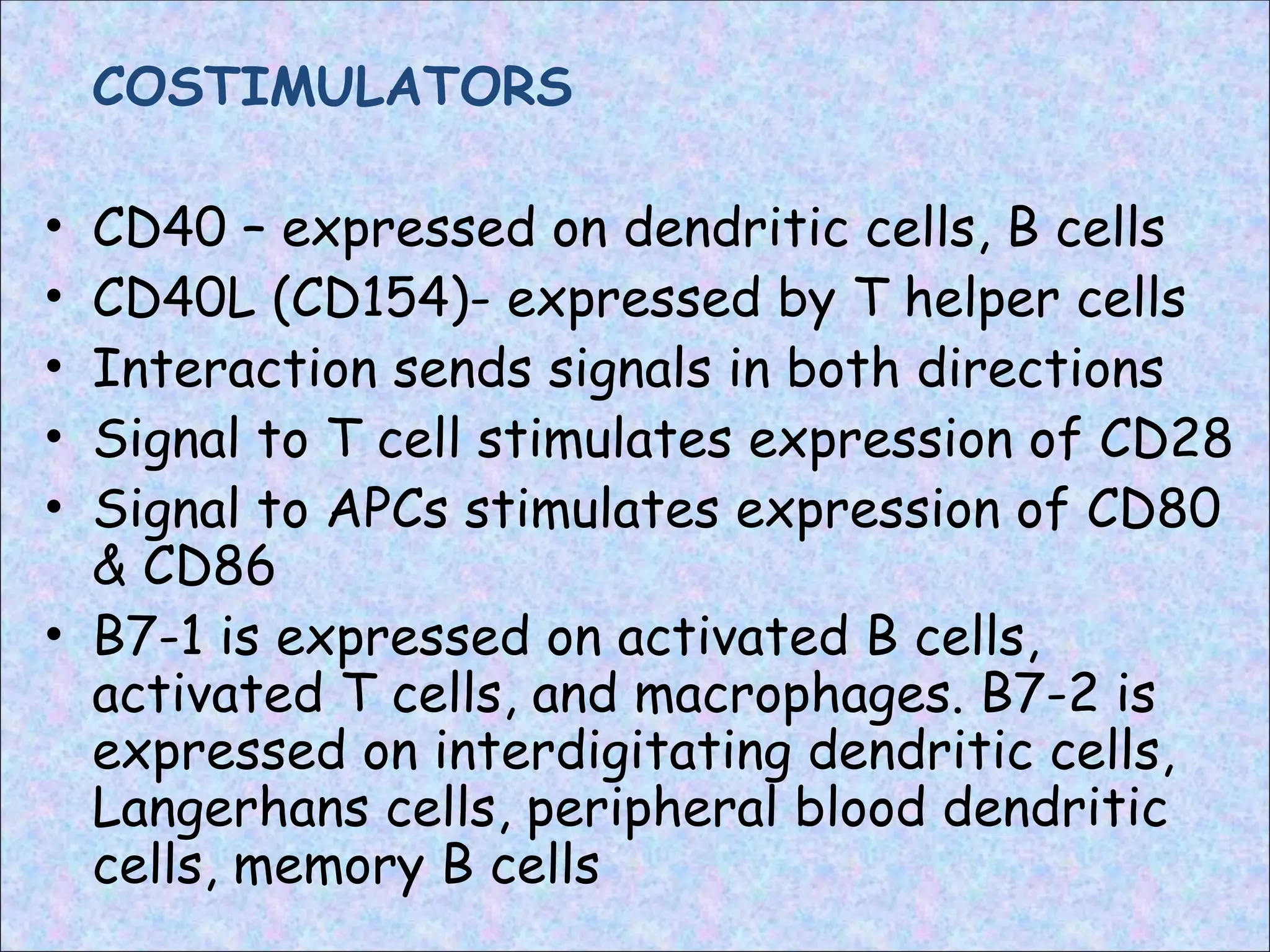
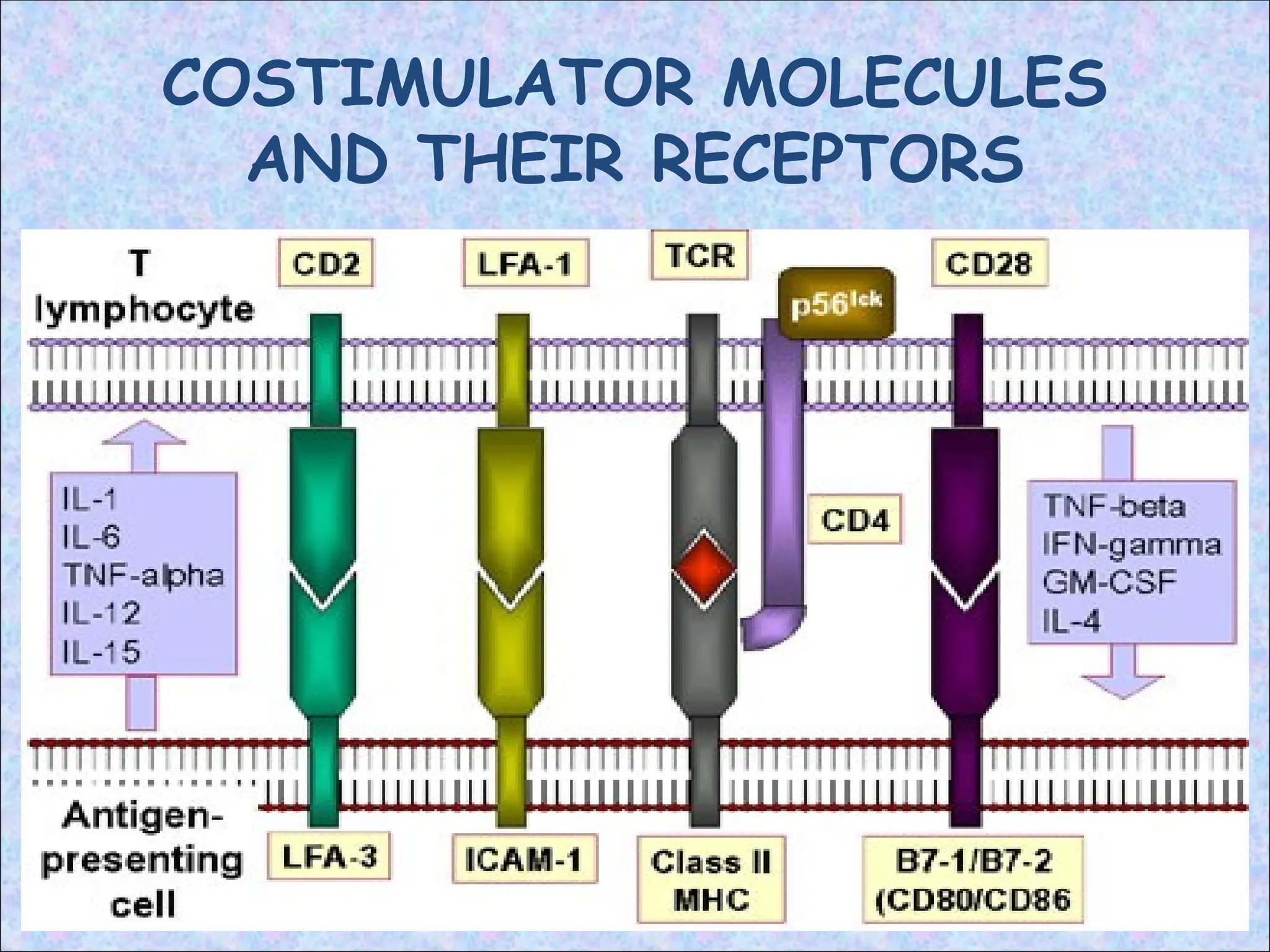
COSTIMULATORS
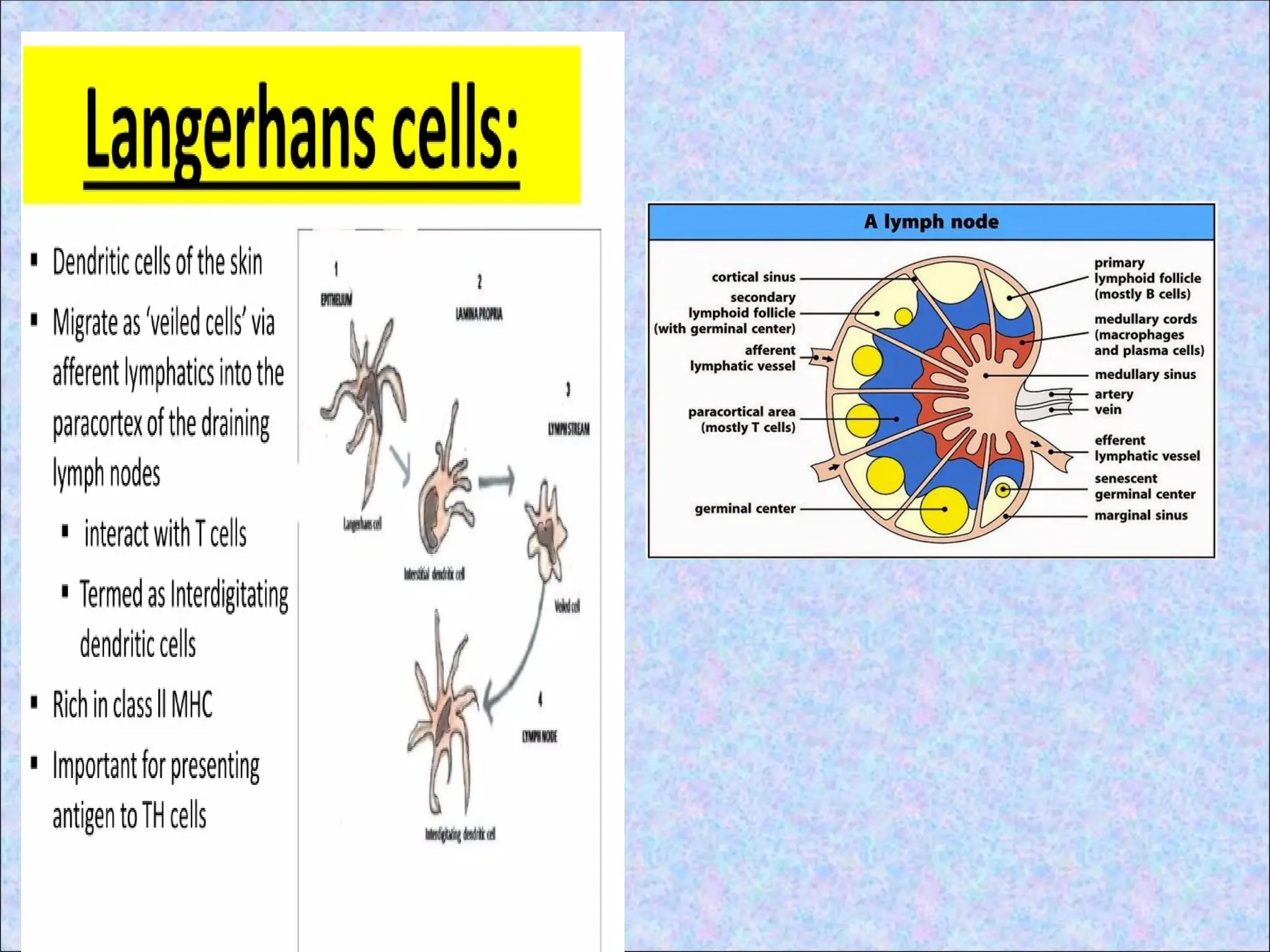
• CD40 –expressed on dendritic cells, B cells
• CD40L (CD154)- expressed by T helper cells
• Interaction sends signals in both directions
• Signal to T cell stimulates expression of CD28
• Signal to APCs stimulates expression of CD80
& CD86
• B7-1 is expressed on activated B cells,
activated T cells, and macrophages. B7-2 is
expressed on interdigitating dendritic cells,
Langerhans cells, peripheral blood dendritic
cells, memory B cells
39.
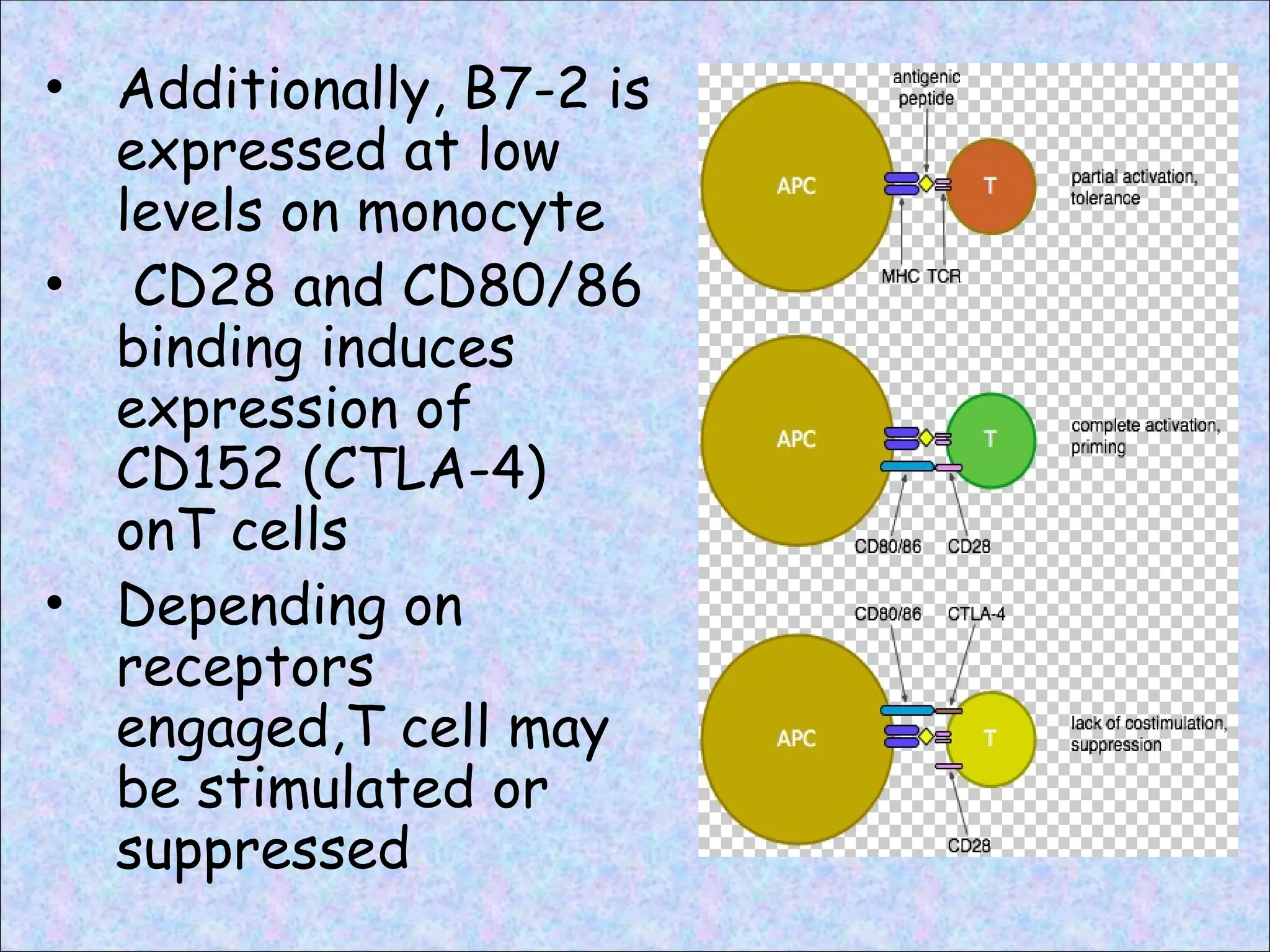
• Additionally, B7-2is
expressed at low
levels on monocyte
• CD28 and CD80/86
binding induces
expression of
CD152 (CTLA-4)
onT cells
• Depending on
receptors
engaged,T cell may
be stimulated or
suppressed
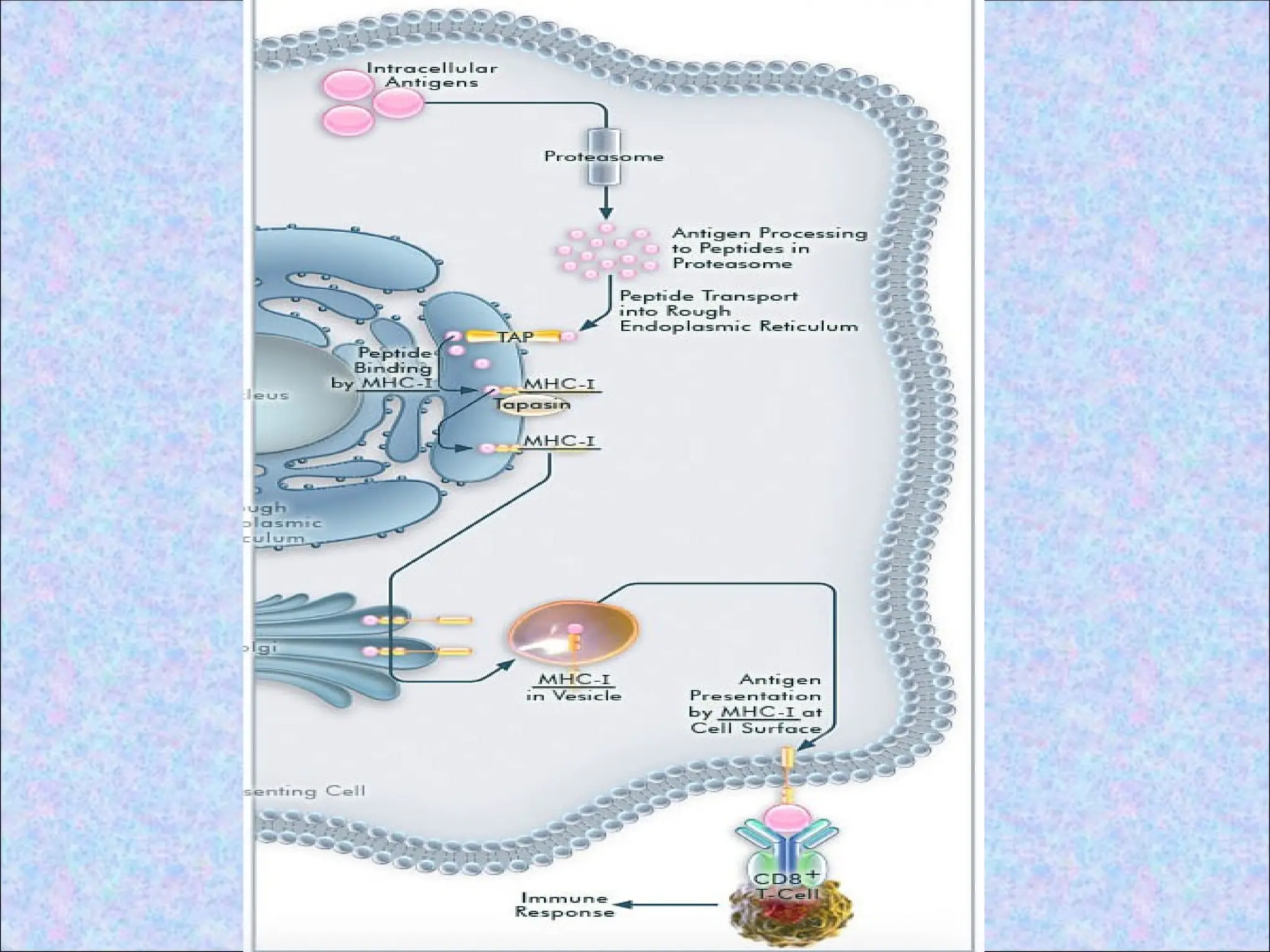
1. Proteolytic degradationof cytosolic
proteins:
• By proteasome
• Proteasome is composed of two inner and
two outer rings, each ring being
composed of seven subunits
4-subunit rings
10 –20 A
43.
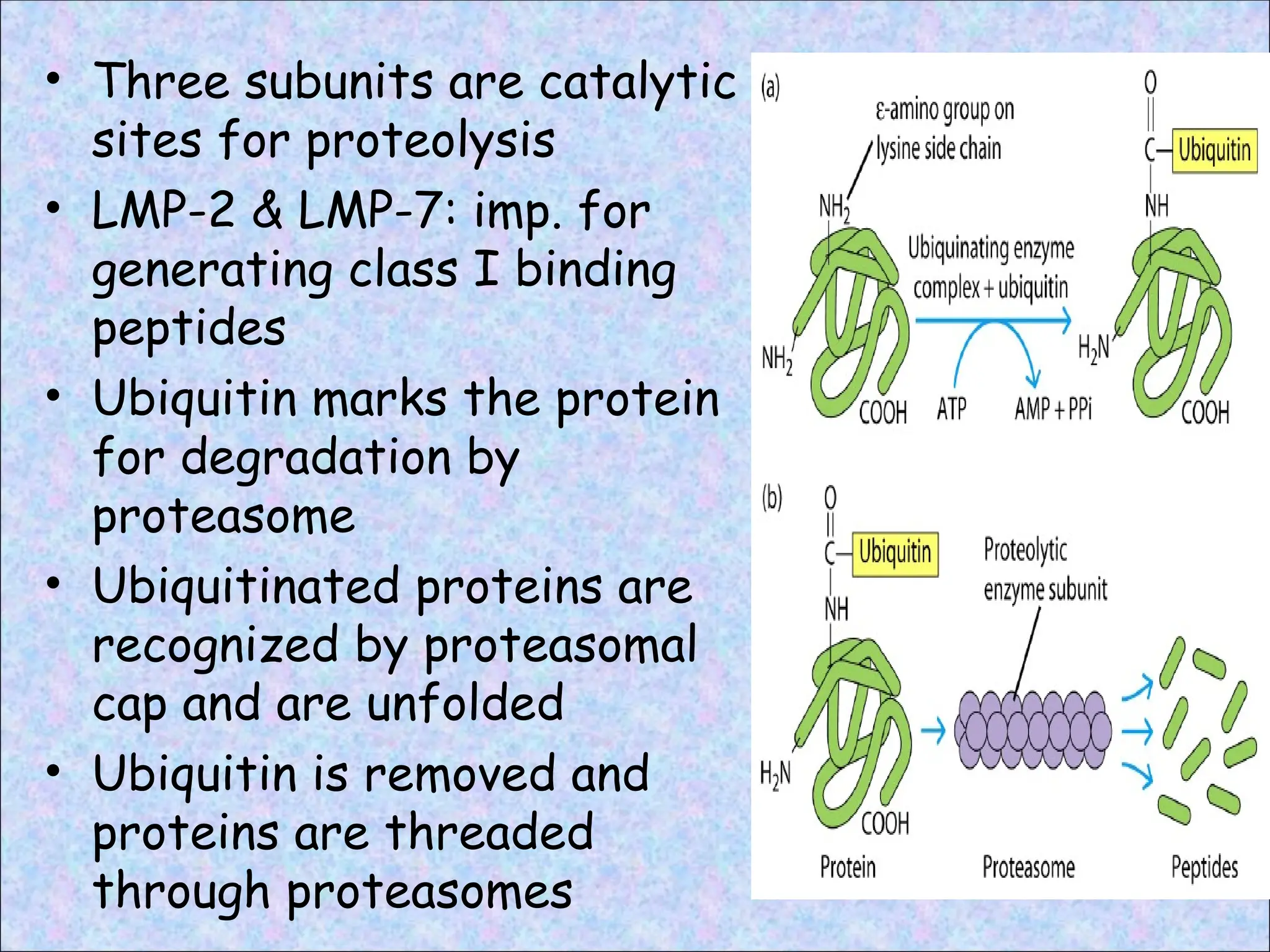
• Three subunitsare catalytic
sites for proteolysis
• LMP-2 & LMP-7: imp. for
generating class I binding
peptides
• Ubiquitin marks the protein
for degradation by
proteasome
• Ubiquitinated proteins are
recognized by proteasomal
cap and are unfolded
• Ubiquitin is removed and
proteins are threaded
through proteasomes
44.
2. Transport ofpeptides from cytosol to the
endoplasmic reticulum:
• Antigenic peptides for class I pathway are
generated in cytosol but newly synthesised
class I MHC molecules are available in the
ER to bind the peptides
• Specialized transporter- TAP
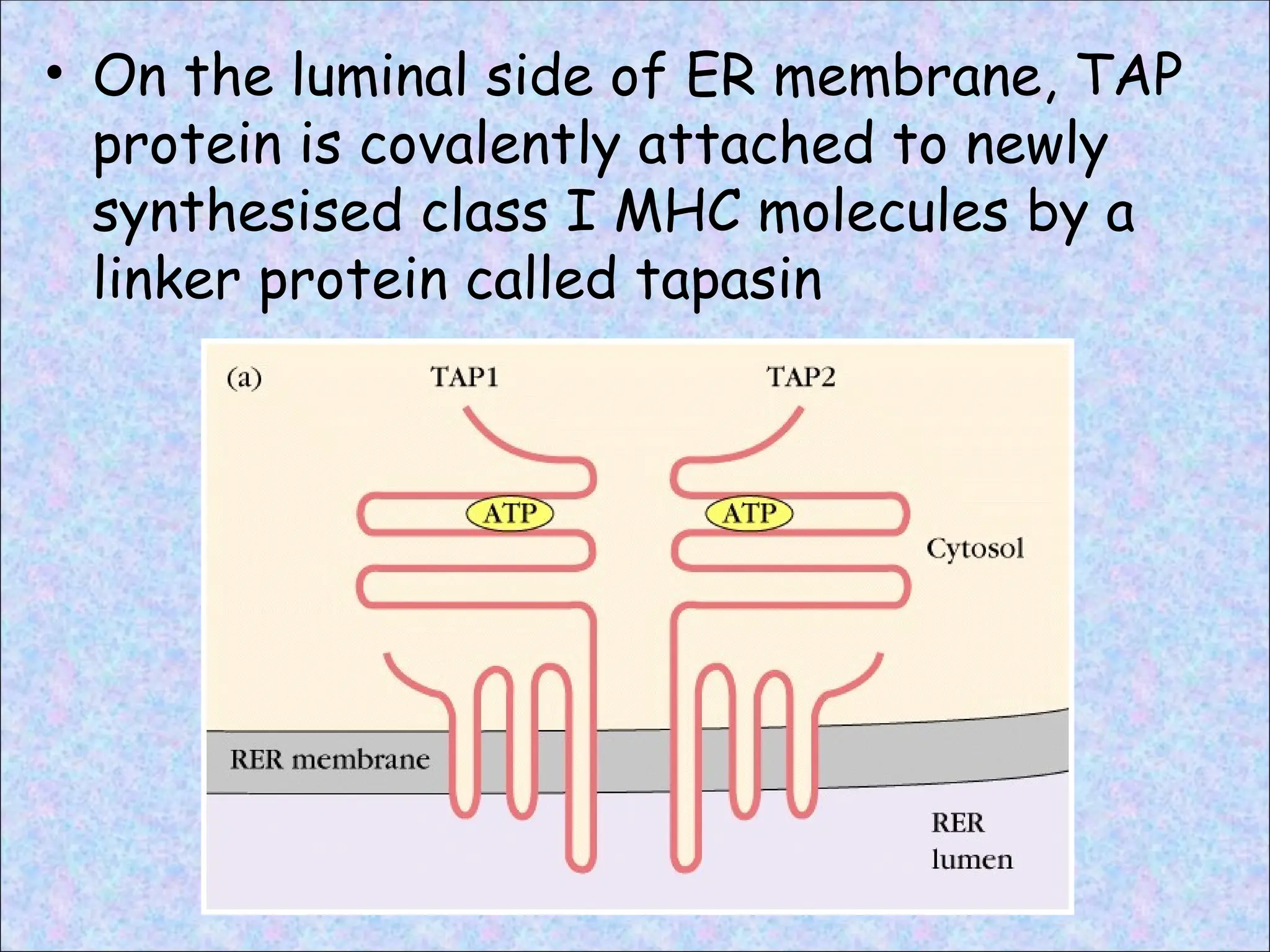
• TAP protein is located in the ER membrane,
where it mediates the active, ATP-
dependent transport of peptides from
cytosol into ER lumen
• TAP transports peptides that are 8 to 13
amino acids long and containing carboxyl
termini that are basic or hydrophobic
45.
• On theluminal side of ER membrane, TAP
protein is covalently attached to newly
synthesised class I MHC molecules by a
linker protein called tapasin
46.
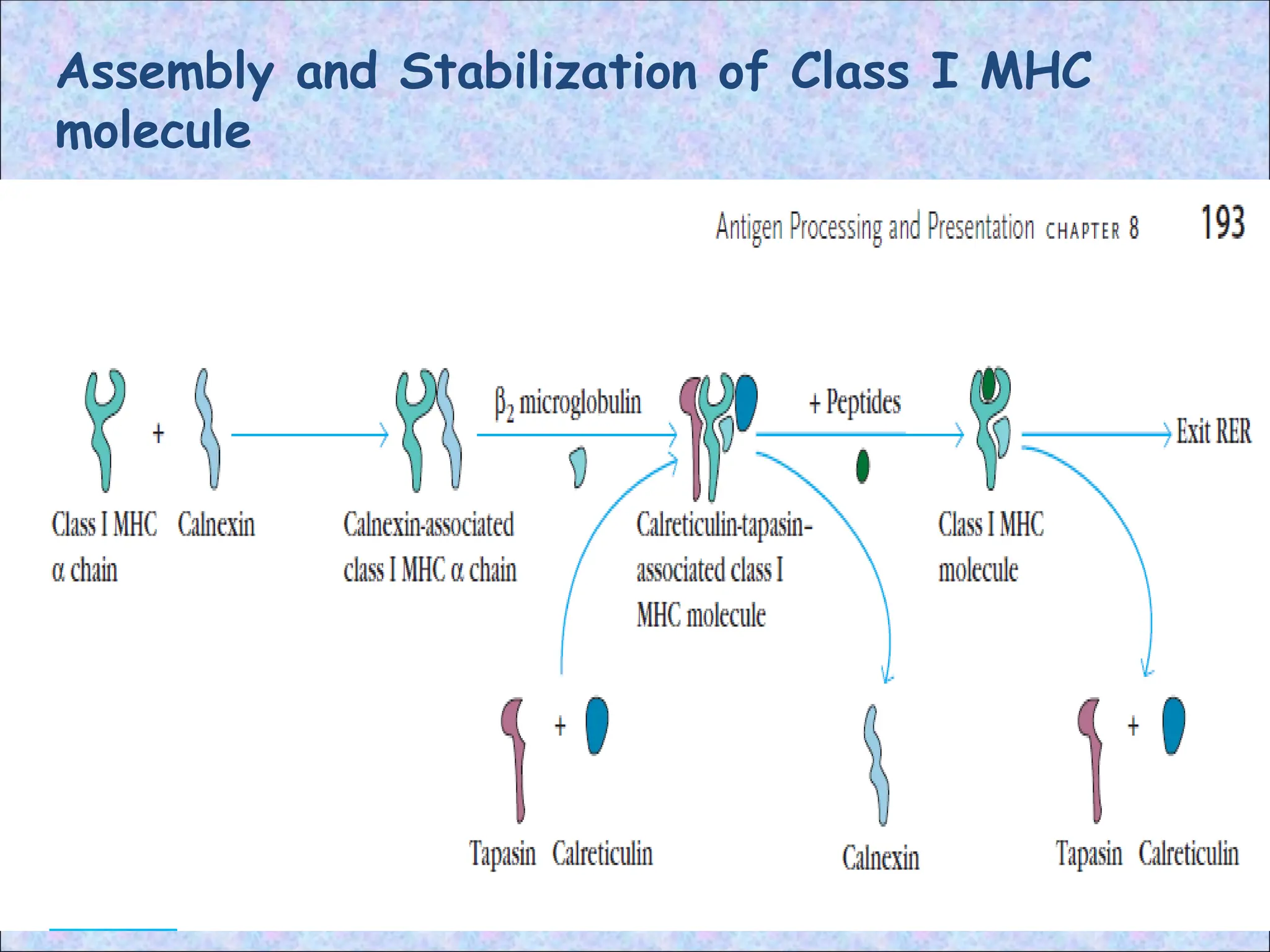
3. Assembly ofpeptide-class I MHC
complexes in the ER:
• Peptides translocated into the ER bind to
class I MHC molecules that are attached
to the TAP dimer
• After entry peptides are trimmed to
appropriate size for MHC binding by
ERAP
• In the absence of bound peptide, many of
the newly formed α chain and β2-
microglobulin dimers are unstable and are
degraded in the ER
47.
• Peptides transportedinto ER bind to class
I, but not class II MHC molecules
because:
• Newly synthesised class I MHC molecules
are attached to the luminal aspect of the
TAP complex, ready to receive peptides
• In the ER, peptide binding clefts of newly
synthesised class II molecules are blocked
by associated Ii
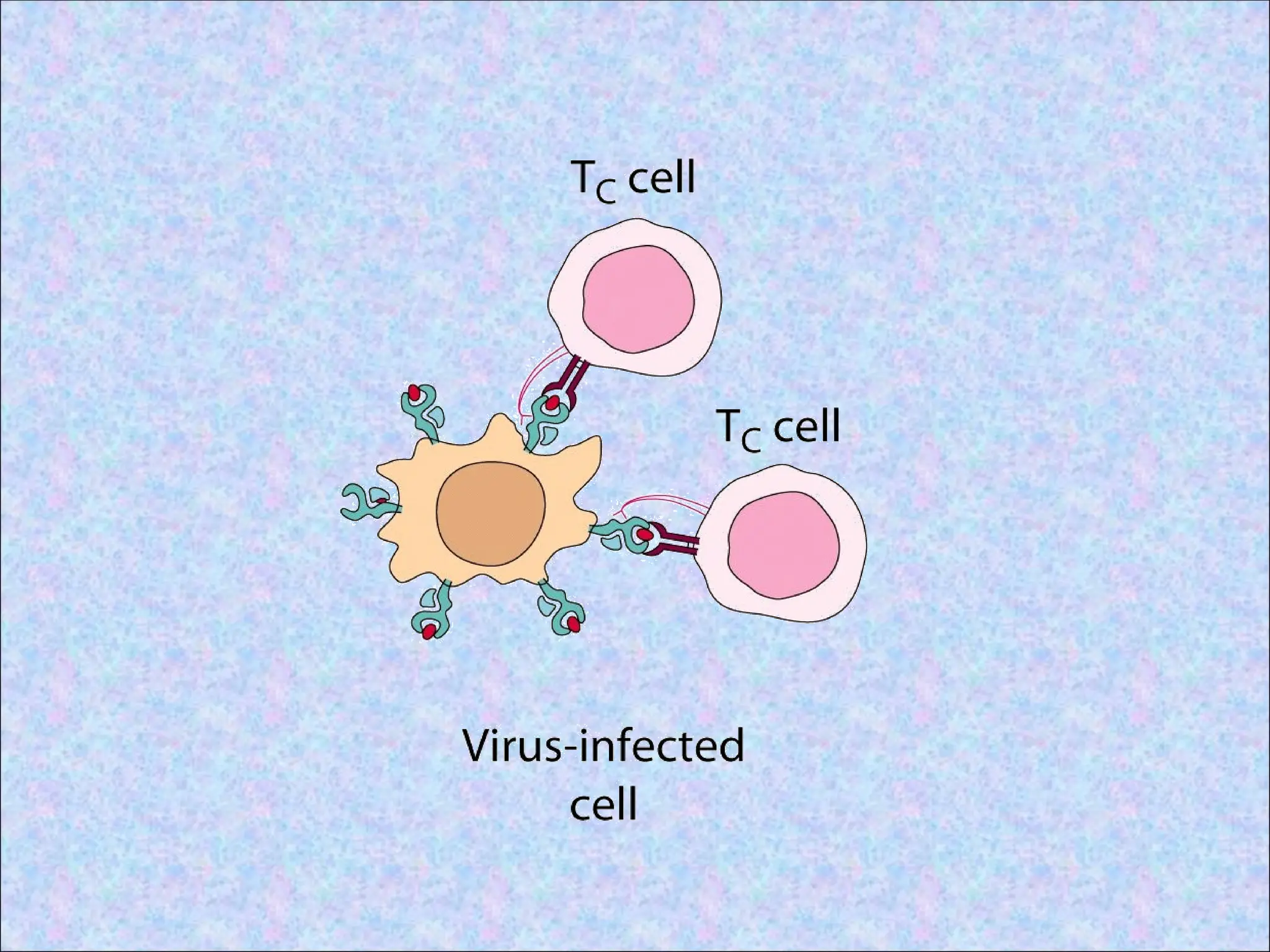
4. Surface expressionof peptide-class I
MHC complexes:
• Class I MHC molecules with bound
peptides are structurally stable and are
expressed on the cell surface
• Stable peptide-class I MHC complexes
move through the Golgi complex and are
transported to the cell surface by
exocytic vesicles
• Recognized by peptide antigen specific
CD8+ T cells
• CD8+ T cells kill the target cell
52.
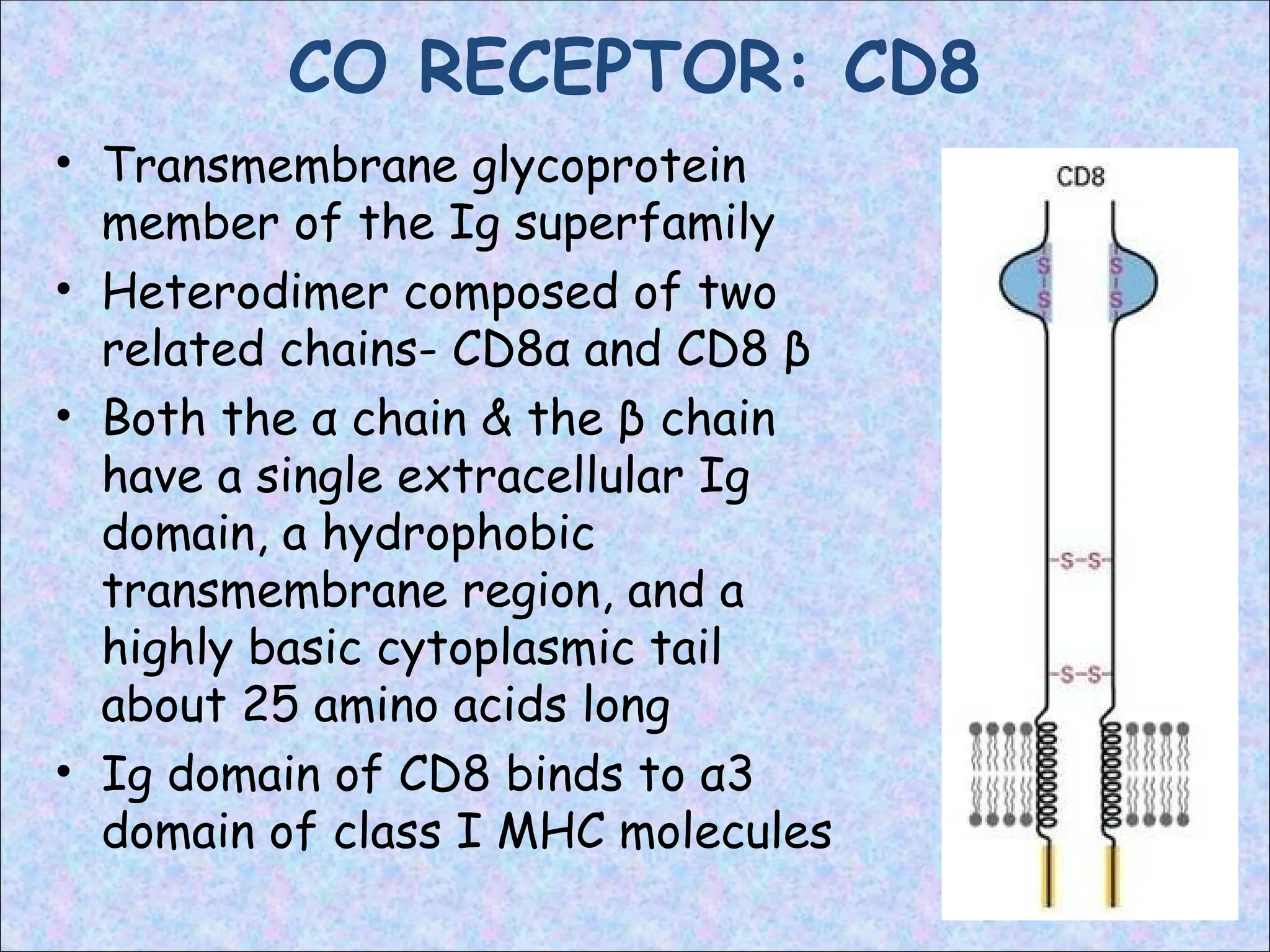
CO RECEPTOR: CD8
•Transmembrane glycoprotein
member of the Ig superfamily
• Heterodimer composed of two
related chains- CD8α and CD8 β
• Both the α chain & the β chain
have a single extracellular Ig
domain, a hydrophobic
transmembrane region, and a
highly basic cytoplasmic tail
about 25 amino acids long
• Ig domain of CD8 binds to α3
domain of class I MHC molecules
54.
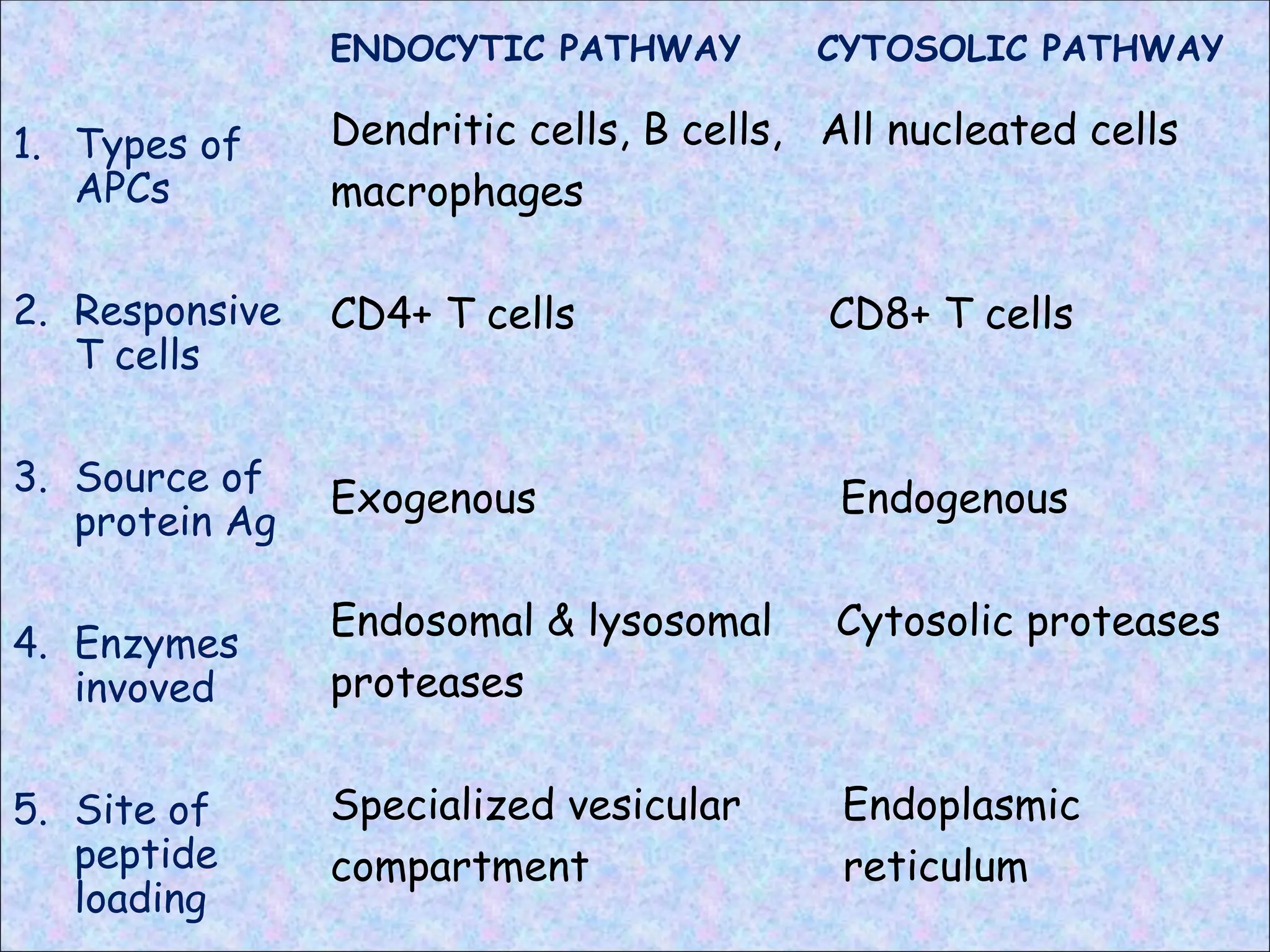
ENDOCYTIC PATHWAY
1. Typesof
APCs
2. Responsive
T cells
3. Source of
protein Ag
4. Enzymes
invoved
5. Site of
peptide
loading
CYTOSOLIC PATHWAY
Dendritic cells, B cells, All nucleated cells
macrophages
CD4+ T cells CD8+ T cells
Exogenous Endogenous
Endosomal & lysosomal Cytosolic proteases
proteases
Specialized vesicular Endoplasmic
compartment reticulum































![ANTIGEN PROCESSING PRESENTATION AND RECOGNITION - Copy [Autosaved].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/antigenprocessingpresentationandrecognition-copyautosaved-220815200136-89a4c3c9-thumbnail.jpg?width=640&height=640&fit=bounds)

















































