1. DYNEIN DYSFUNCTION IMPAIRS MUSK CLUSTERING AT THE NEUROMUSCULAR JUNCTION:
IMPLICATION FOR NEURODEGENERATION IN ALS
Background
A great deal is known about neuromuscular junction (NMJ) formation but much less is
known on how they are stabilized and maintained. Loss of NMJ integrity occurs in
motor neuron diseases such as amyotrophic lateral sclerosis (ALS). Cytoplasmic
dynein, together with its activator dynactin, forms a large motor protein complex
responsible for the majority of microtubule‐mediated minus‐end transport of organelles.
Studies on mouse models expressing mutations in dynein have shown mild
neurodegeneration linked to defective retrograde transport in neurons and NMJ
defects. Many pieces of evidence have (re) placed the muscle as one of initiating
centers of ALS and because dynein complex has been described as important
protagonists of muscle development, a key question is whether the motor protein
complex can play a role in muscle‐derived pathobiology in ALS.
Objective
To investigate the role of the dynein in the formation and maintenance of the post-synaptic
apparatus and the underlying stabilizing mechanisms.
Discussion & Conclusions
Dynein is important for the maintenance of the post-synaptic apparatus via control of MuSK localization,
trafficking and clustering at the neuromuscular junction.
These preliminary data secure our in-vitro differentiation protocol as a robust model to study highly
differentiated fibers recapitulating muscle development in SOD1G93A mice.
Our findings from SOD1G93A mice revealed that features of the post-synaptic apparatus are impaired in fibers
taken from young mice which are still asymptomatic supporting the hypothesis that muscle development could
be an independent trigger of neuromuscular junction loss and muscle atrophy.
This novel model could be used as a new tool to study time-based development of neuromuscular structures
in SOD1G93A mice and to test new therapeutic compounds for ALS.
Valérie VILMONT1, Bruno Cadot1 and Edgar R. Gomes1,2
1Myology Research Center, Sorbonne Universités, UPMC Université Paris, Paris, France, 2Instituto di Medicina Molecular, Lisbon, Portugal
Email addresses for correspondence: vilmont@myologygroup.net, er.gomes@gmail.com
Keywords: Dynein, NMJ, Muscle-specific tyrosine kinase, endosomal trafficking, SOD1G93A
A) In vitro models
1-Primary myoblasts were isolated from P7-WT littermates or transgenic pups expressing the
G93A mutant form of human SOD1. Human SOD1 is known to aggregate dynein causing loss
of dynein function..
2-Myoblasts at 70-80% confluence were directed to differentiate to Day 6 (middle point
differentiation) and Day 9 (end point differentiation) on Matrigel support2 in the presence
of agrin to induce AchR clustering which are considered as surrogate NMJ.
3-Myofibers were treated with either small interfering RNAs against dynein (shDHC) or
ciliobrevinD to impair dynein motor function.
4- Post-synaptic features were assessed qualitatively and quantitatively at Day 6 and Day 9 in
conditions cited in 3-
B) Ex vivo model
1- Extensor digitorum longus (EDL) single fibers were isolated from symptomatic SOD1G93A
and littermates and postsynaptic features were assessed.
REFERENCE
1Dobrowolny, JCB, 2005; Dupuis, Exp Neu 2009; Wong and Martin Hum Mol Genet, 2010
2Falcone S et al., EMBO Mol Med, 2014
3http://neuromuscular.wustl.edu/synmg.html
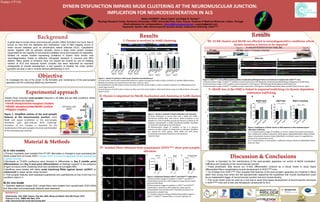
Figure 2. Simplified scheme of the post-synaptic
features at the neuromuscular junction. AchR,
MuSK and rapsyn localization at the post-synaptic
membrane upon agrin-induced AchR clustering3.
Formation of this complex is important for the
maintenance of the post-synaptic structure and thereafter
of the neuromuscular junction.
Figure 4. Comparison between SOD1WT and SOD1G93A single fibers.
A) Representative images of myofibers in SOD1WT and SOD1G93A
isolated fibers stained for α-BTX (magenta), rapsyn (green), MuSK
(red) and DAPI (blue).
B) Representative images of myofibers in SOD1WT and SOD1G93A
isolated fibers stained for α-BTX (magenta), rapsyn (green),
phosphoMuSK (red) and DAPI (blue). Myofiber boundaries are
represented as dashed lines.
C, D, E) Mean intensity of Rapsyn/MuSK/PhosphoMuSK signal in
SOD1WT vs SOD1G93A isolated fibers. Error bars represent sd.
Results
I
SOD1 G93A MOUSE LIFESPAN
ASYMPTOMATIC
P7 pups
SYMPTOMATIC
=30 days =120days
SOD1G93A
Day6
Day9
SOD1G93ASOD1 WT
Figure 1. Dynein is involved in AchR cluster formation and maintenance
A) AchR clusters stained with tetramethylrhodamine-coupled α-Bungarotoxin (BTX) at Day 6 and Day 9 of myofiber differentiation.
Myofiber boundaries are represented as dash lines. Scale bar 10µm.
B) Quantification of AchR clusters number per fiber and AchR cluster length in control vs shDHC conditions at 2 differentiation time
points (days 6 and 9).
C) Quantification of AchR clusters number per fiber and AchR cluster length in ciliobrevinD release assay at 2 differentiation time points
(days 6 and 9).
A
B C
II- Dynein is important for MuSK localization and clustering at AchR clusters
III- AChR clusters and MuSK are affected in neurodegenerative conditions where
dynein function is known to be impaired
Figure 2. Dynein is involved in MuSK localization and clustering
A) MuSK localization in control (fiber with a diese) and shDHC-
transfected myofiber (fiber with asterix). MuSK localization at cell
periphery in control myofiber (arrows) and absence of MuSK at cell
periphery (arrowheads) in shDHC-transfected myofiber. Myofibers
are stained for MuSK (grey) and DAPI (blue).
B) Representative images of myofibers at Day 6 in absence,
presence or after release of ciliobrevinD as indicated. Myofibers
are stained for α-BTX (green), MuSK (Red) and DAPI (blue).
Arrowheads show MuSK localization at cell periphery.
Scale bar 10µm.
Figure 3. SOD1G93A in vitro differentiated myofibers recapitulate pathological features reminiscent of symptomatic SOD1G93A mice.
A) Representative images of SOD1G93A myofibers at Day 6 stained for α-BTX (Magenta), MuSK/phosphoMuSK/Rapsyn (Magenta) and DAPI (blue).
B) Representative images of WT myofibers at Day 9 stained for α-BTX (green), MuSK/phosphoMuSK (Red) and DAPI (blue) and
C) Representative images of SOD1G93A myofibers at Day 9 stained for α-BTX (green), MuSK/PhosphoMuSK (Red) and DAPI (blue). Scale bar 10µm.
A
B C
IV- Isolated fibers obtained from symptomatic SOD1G93A show post-synaptic
alteration
V- MuSK loss at the NMJ is linked to impaired trafficking via dynein-dependent
endosomes trafficking
Figure 5. Stabilization of MuSK clusters at NMJ depends on Rab-mediated dynein
dependent trafficking.
A, C, E) Representative images of myofibers in control, CiliobrevinD treated and released
from ciliobrevinD conditions stained for MuSK (green), Rab4a/Rab7/Rab11 (Red) and DAPI
(blue). Colocalization between Rab4a /Rab7/Rab11 and MuSK clusters is magnified (2x).
Arrows show colocalization points.
B, D, F) Quantification of colocalization coefficients between MuSK and Rab4a/Rab7/Rab11.
Scale bar 10µm.Material & Methods
Experimental approach
In vitro DIFFERENTIATION TIMELINE
Assess three important post-synaptic features in in vitro and ex vivo conditions where
dynein functions are impaired.
AchR (Acetylcholine receptor) clusters
MuSK/ phosphorylated MuSK clusters
Rapsyn clusters
Results
II- Dynein is involved in AchR clustering
Poster n°P116
scramble
shDHC
scramble
shDHC
Rab4=early Rab7=late Rab11=recycling
B D F
Day 9=Day 6=