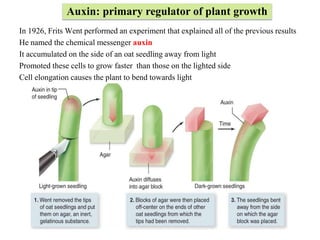
1. In 1926, Frits Went performed an experiment that explained all of the previous results
He named the chemical messenger auxin
It accumulated on the side of an oat seedling away from light
Promoted these cells to grow faster than those on the lighted side
Cell elongation causes the plant to bend towards light
Auxin: primary regulator of plant growth
2. IAA biosynthesis is associated with rapidly growing tissues mainly in
SAM, young leaves, fruits etc.
Tryp IPyA IAATAA YUC
Synthesized from tryptophan via IPyA or acetonitrile as intermediates,
Auxin can be conjugated with amino acids or hexose sugars leading to
inactivation and degradation to maintain homeostasis
Physiological function and cellular level responses
Promotes growth in stems and coleoptiles while inhibiting growth in
roots maintaining apical dominance
Organogenesis, vascular tissue differentiation
Auxin induces cell wall creep and cell elongation by proton extrusion
auxin induces the activity of plasmamembrane H+ ATPases, acid induced
growth renders plant wall for long term extension called creep (time
dependent long term extension). Expansin protein also causes loosening of
cell wall
Phototropism, gravitropism determined by lateral distribution of auxin
During gravitropism the tip of the coleoptile could perceive gravity and
redistribute auxin to the lower side
Auxin biosynthesis and function
4. Starch statolith concept and polar auxin transport
During plant reorientation, a plant is
rotated relative to the gravity vector. This
results in the sedimentation of dense
amyloplasts within the statocytes. In roots
the statocytes are the columella cells,
whereas in stems they are the endodermal
cells. Each endodermal cell contains a
large vacuole, and the amyloplasts must
traverse it by tunneling through trans-
vacuolar strands in order to reach the new
lower side of the cell. This requires proper
vacuole structure, which the SGR proteins
mediate.
Amyloplast sedimentation then activates
signal transduction through second
messengers, possibly calcium ions or
protons, or InsP3. In a process that is not
completely understood, the second
messengers activate the relocalization of
auxin transporters, such as PIN3 and PIN7
in the columella cells. The new polarized
distribution of auxin efflux carriers
changes the flow of auxin through out the
plant.This differential auxin transport
affects cell elongation rates,there by
resulting in organ curvature as the plant
grows.
5. Polar auxin transport
In the acidic apoplastic environment, a
fraction of the weak acid IAA exists in its
undissociated form, which can passively
diffuse through the plasma membrane inside
the cell. By contrast, the non-lipophilic and
therefore less permeable proton-dissociated
auxin fraction requires the amino acid
permease-like AUX1, which catalyzes proton
symport activity, to enter the cell. In the
neutral cytosolic environment, IAA exists
mainly in its membrane-impermeant anionic
form that requires active transport to exit the
cell.
The best characterized auxin efflux carriers
are members of the unique and plant-specific
PIN protein family, believed to be secondary
transporters energized by proton gradients. By
contrast, some plant homologs of the human
MDR/PGP transporters belonging to the
ABCB subfamily, such as ABCB1, ABCB4,
and ABCB19, have been implicated in ATP-
energized auxin efflux.
6. Positive Negative
Primary interaction
Secondary interaction
Two input pathways
regulate separate
responses
Two input pathways
regulate a single
shared protein
Tertiary interactions
One of the signaling
pathway regulates
other
Plant Signaling: crosstalk between different signaling molecules
8. The core components of the auxin signaling machinery belong to three
protein families:
F-box TRANSPORT INHIBITOR RESPONSE 1/AUXIN SIGNALING F-BOX
PROTEIN (TIR1/AFB) auxin co-receptors
Auxin/INDOLE-3-ACETIC ACID (Aux/IAA) transcriptional repressors
AUXIN RESPONSE FACTOR (ARF) transcription factors
Aux/IAAs comprise three functional domains:
Leucine repeat EAR motif within domain I,
Internal domain II that contains a GWPP-core degron motif,
C-terminal region that forms a type I/II Phox and Bem1 (PB1)
domain
The PB1 domain facilitates interactions with ARF proteins as well as self-
dimerization ,while the degron motif is required for interaction with
TIR1/AFB proteins and therefore determines Aux/IAA stability. Domain I
functions as a repression motif by recruiting transcriptional co-repressors.
Although domain II is necessary for the interaction with TIR1/AFBs,
other sequences outside of domain II, including an N-terminal lysine-
arginine (KR) motif, contribute to TIR1/AFB binding and/or Aux/IAA
degradation
Auxin signaling
10. A: GA-dependent GID1-DELLA complex formation resulting in DELLA recognition
and ubiquitylation by the SCFSLY1 E3. Polyubiquitylation leads to DELLA proteolysis by
the 26S proteasome, thereby lifting DELLA repression of GA responses.
B: Proteolysis-independent GA signaling in sly1 mutants occurs when GID1-GA-DELLA
complex formation blocks DELLA repression of GA responses without DELLA destruction.
C: EL1-mediated phosphorylation of DELLA activates DELLA as a repressor of GA
responses
GA signaling
11. In the night, GA and GID1 levels are
highest, thus repressing DELLA’s
ability to negatively regulate PIF. PIF
mRNA levels rise during the night,
leading to rapid hypocotyl growth
before morning. At dawn, a drop in
GID1 levels results in DELLA
stabilization and repression of PIF,
leading to greening and slower
growth. In the day, red light actives
Pfr, which phosphorylates PIF,
leading to PIF destruction via the
ubiquitin-proteasome pathway. Lack
of PIF blocks hypocotyl growth.
DELLA modulation of PIF activity
16. Growth or Defense: Who will win? (1) Jasmonate-induced defense responses
reduce carbon availability by diverting
resources to the production of specialized
metabolites. Altered carbon status may be
detected by energy-sensing systems to reduce
growth rate to a level that matches carbon
availability. (2) MYC transcription factors
inhibit growth independently of jasmonate-
induced defense responses. In roots, for
example, MYC2 decreases the activity of the
root meristem by repressing the expression of
AP2-domain PLETHORA (PLT)
transcription factors. In leaves, MYCs
repress the expression of photosynthesis
genes and may also constrain growth through
unidentified regulators that repress cell
division or cell expansion. (3) Crosstalk
between JA-Ile and GA signaling pathways
modulates growth responses. JAZ
degradation releases DELLA proteins, which
negatively regulate PIF transcription factors
to inhibit cell expansion-type growth. PIF
transcription factors inhibit chlorophyll
biosynthesis and stimulate auxin production
as part of the shade avoidance growth
response
17. Ethylene (C2H4) is a gaseous hormone with
diverse actions
Ethylene regulates:
•fruit ripening
•organ expansion
•senescence
•gene expression
•stress responses
Cotton plants
7 days ethyleneAir (control)
Air Ethylene
Arabidopsis
Beyer, Jr., E.M. (1976) A potent inhibitor of ethylene action in plants. Plant Physiol. 58: 268-271.
18. Early fruit-ripening practices
Ethylene in smoke has long
been used to ripen fruit; this
practice has included
ripening pears in the smoke
from incense. Gashing of
unpollinated figs has also
been practiced; the ethylene
produced upon wounding
induces ripening
Image sources: British Museum; Kurt Stüber
19. When germinating in the dark, impeded
seedlings produce ethylene which confers a
characteristic “triple response”
C2H4
C2H4
Ethylene induces the
triple response:
•reduced elongation,
•hypocotyl swelling,
•apical hook
exaggeration
It’s thought that this
response helps the
seedling push past the
impediment
By treating dark-grown seedlings with exogenous
ethylene, ethylene-response mutants could be
identified quickly and easily based on the triple
response phenotype
20. Ethylene’s roles in whole-plant processes
• Shoot and root elongation
• Reproductive development
• Sex determination
• Petal and leaf senescence
• Fruit ripening
• Flooding responses –
• Aerenchyma formation, leaf epinasty
• Deepwater rice
• Pathogen responses
• Interactions with other hormones
21. The two key enzymes, ACS and ACO, are rare and
unstable
ACS is ACC synthase
ACO is ACC oxidase
Both are unstable
proteins. Normally ACS
is continually
synthesized and
continually degraded,
maintaining a very low
level of ethylene
Stress-induced protein
phosphorylation stabilizes
ACS and increases
ethylene accumulationACS
WOUNDING,
STRESS,
PATHOGEN
ATTACK
P
Proteasome
23. Ethylene synthesis and homeostasis - summary
Simple biosynthetic pathway regulated by expression and
stability of ACS and ACO
ACS and ACO activities are tightly regulated transcriptionally
and post-transcriptionally and sensitive to developmental cues,
wounding and pathogen attack
Ethylene
Biosynthesis
SAM
ACC
C2H4
ACS
ACO
ACS proteins
stabilized by
wounding, other
hormones
25. ETHYLENE RESPONSE1 (ETR1) encodes an
ethylene receptor
ETR1 was the first protein to be unambiguously
identified as a phytohormone receptor (1993)
ETR1 binds ethylene
ETR1 is similar in sequence to known-receptors in animal
cells
ETR1 is membrane localized
ETR1
histidine kinase receiverGAF
ethylene
binding
Ethylene-binding,
membrane-spanning domain
Arabidopsis has
five ethylene
receptors, with
differing signaling
strengths
26. The etr1-1 mutation is dominant
WT etr1-1WT WT WT
etr1-1etr1-1ETR1
Introduction of the
mutant etr1-1 allele into a
wild-type plant causes an
ethylene insensitive
phenotype
27. How can a mutant receptor have a dominant
phenotype???
Responses
ON
Ethylene
Responses
OFF
The receptors negatively regulate the responses
No Ethylene
When not
bound to
ethylene, the
receptor
shuts off the
ethylene
response
When bound
to ethylene,
the receptor
does not
shut off the
ethylene
response
31. • By increasing the cytokinin concentration in the shoot, lateral buds become active
resulting in increased branching (and a more bushy shoot development)
Cytokinin
Cytokinin and shoot apical dominance
32. The effect of cytokinin on
senescence.
Cytokinin applied to the right-
hand primary leaf of this bean
seedling inhibited its
senescence. The left-hand did
not get cytokinin.
Richmond Lang effect
33. Cytokinin signaling in plants
The cytokinin receptors histidine kinases are primarily
localized on the endoplasmic reticulum, as well as on the
plasma membrane. Cytokinin binds to AHK proteins,
inducing conformational changes that trigger a
phosphorelay. A phosphoryl group (P) is first transferred
from a conserved His (H) to an Asp (D) residue within the
receptor and is then relayed to five histidine
phosphotransferase proteins (AHP1-AHP5). The pseudo-
HP AHP6 inhibits cytokinin signalling by competing with
AHP1-5 for phosphotransfer. The AHPs continuously
translocate between the cytosol and the nucleus, where
the Arabidopsis response regulators (ARRs) are in turn
phosphorylated. Phosphorylation of the type A ARRs
stabilizes them. The phosphorylated type B ARRs can
bind DNA and initiate transcription of cytokinin-
responsive genes, including the type A ARRs, which act
as inhibitors of cytokinin signalling.
34. ABA : stress regulator
When the plants are exposed to osmotic
stress, accumulating ABA binds to the central
hydrophobic moiety of PYL receptor leading
to a conformational change which then
induces the binding of PP2Cs to form a
stable close PYL-ABA-PP2C complex. The
protein phosphatase activity of PP2C is
inhibited by ABA-PYL complex and this
activates SNF1 related protein kinase 2
(SnRK2) protein by autophosphorylation.
The autophosphorylation of SnRK2s switch
on the downstream mitogen activated protein
kinase (MAPK) signalling cascade
35. ABA, during stomatal
closure activates CPKs,
OST1 as well as RBoH
protein to depolarize the
membrane of Guard cell
by regulating different
anion channels
Role of ABA during stomata closure
36. ABA signalling during stress responses
During stomatal closure, ABA-activated SnRK2 also induces phosphorylation of the
plasma membrane NADPH oxidase, respiratory burst of oxidase homolog F (RbohF),
generating O2
- and subsequently H2O2 within the apoplastic region of guard cells. The
apoplastic ROS then enters into the cell via plasmamembrane intrinsic protein 2.1 (PIP2.1)
and activates two signalling components, guard cell hydrogen peroxide-resistant 1 (GHR1)
and S-type anion channel (SLAC1), which aggravate plasma membrane calcium (Ca2+)
channels. The elevation of Ca2+ activate Ca2+ dependent protein kinases like calcium
dependent protein kinase (CPKs) and calcineurin B like protein1/9 (CBL1/9) – CBL
interacting protein kinase 26 (CIPK26) for further phosphorylation of RbohF.
Ca2+ mediated and NO mediated