1. Analysis of Alzheimer disease pathology and Nurr1 in AD mice
Sarah Metcalfe2; Rebecca Courtney1; Gary E. Landreth1
1Alzheimer’s Disease Research Laboratory, Department of Neuroscience, Case Western Reserve University, Cleveland, OH 44106
2Department of Biology, St. Bonaventure University, St. Bonaventure, NY 14778
ABSTRACT
Alzheimer disease (AD) is a neurodegenerative disease characterized by neuronal loss,
impaired cognitive function, memory loss, and personality changes. AD pathogenesis includes
the development of plaques formed from amyloid-β (Aβ) protein and neurofibrillary tangles
composed of intracellular tau protein . The accumulation of plaques correlates with increased
brain inflammation as shown by activation of microglia and astrocytes. One of the first brain
areas to exhibit Alzheimer pathology is the hippocampus, which plays a critical role in long-
term memory consolidation and experiences profound neuronal death during the progression
of AD. Hippocampal-dependent memory formation is facilitated by the NR4A subfamily of
nuclear receptors which have broad neuroprotective roles (Hawk and Abel 2011). The NR4A
receptors have been found to be dysregulated in the brains of AD patients and mice with
amyloid pathology (Skerrett et al. 2014), but the time course and cellular localization of these
changes in expression remains largely unstudied.
To study the relationship between NR4A expression and neuronal death in AD we utilized the
B6-5XFAD mouse model, which experiences severe amyloidosis and neuronal death. Our
study focused on the subiculum, part of the main output pathway from the hippocampus to
the cortex, where we observed neuronal death to occur by 4 months of age. We observed
amyloid plaques and microgliosis in the subiculum of B6-5XFAD mice and saw an increase in
plaque area with age. We focused on the Nurr1 (NR4A2) nuclear receptor which has altered
expression with pathology progression in AD mice and has been shown to play a
neuroprotective role. We saw Nurr1 colocalized with neurons in the subiculum, and
determined that overall hippocampal Nurr1 transcript and protein is decreased in B6-5XFAD
mice by 4 months of age. Our study indicates that genetic manipulation of Nurr1 using viral
vectors will be useful if targeted to neurons in mice younger than 4 months of age. A long-
term goal of our study is to both overexpress and knock down neuronal levels of Nurr1 in the
subiculum, to determine its value as a potential novel therapeutic target.
INTRODUCTION
• Plaques are caused by aggregation of extracellular Aβ leading to the formation of
deposits in the brain. Aβ is formed through specific processing of the
transmembrane amyloid precursor protein (APP) and has been shown to have
neurotoxic properties disrupting crosstalk between neurons.
• The 5XFAD mouse model has 5 mutations expressed in neurons, including 3 in APP
and 2 in PS1 (a catalytic subunit of one APP secretase). This AD model produces
amyloid deposits at an earlier age than other models, and these mice experience
neuronal loss and hippocampal based memory impairments.
• The hippocampus is part of the limbic system and involved with memory
formation and storage. Its function includes consolidation of new memories,
emotional responses, navigation, and spatial orientation. The subiculum is the
main output for the hippocampus.
• NR4A receptors, especially Nurr1, are important for hippocampal function and
have neuroprotective roles.
• We explore Nurr1 expression in the hippocampus and in specific cell type in
5XFAD mice. Along with this we look at inflammation, plaque formation, and
neuronal loss in the hippocampus to determine the relationship between Nurr1
expression and AD pathology.
REFERENCES & ACKNOWLEDGMENTS
• Hawk, J. D., & Abel, T. (2011). Brain Research Bulletin, 85, 21-29. http://dx.doi.org/10.1016/j.
brainresbull.2011.02.001
• Saijo, K. et al. (2009). Cell, 137(1), 47-59.http://dx.doi.org/10.1016/j.cell.2009.01.038
• Skerret, R., Malm, T., & Landreth, G. (2014). Neurobiology of Disease, 72, 104116.http://
dx.doi.org/10.1016/j.nbd.2014.05.019
Thank you to SURP for the opportunity to have this experience. Thank you also to the
Landreth lab for allowing me to work alongside them. This work was funded by the
NIH R01 AG030482 and NIH Grant F31 AG 046055.
RESULTS RESULTS
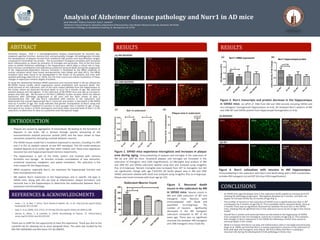
CONCLUSIONS
• As 5XFAD mice age the plaque area in the subiculum tends toward an increase (p=0.07)
showing AD pathology progression. Microgliosis develops by 4 months, and does not
appear to increase further by 10 months of age (Fig 1).
• The number of neurons in the subiculum of 5XFAD mice is significantly less than in WT
counterparts by 4 months of age (Fig 2). This is probably due to neuronal death, since at
2 months There was no significant neuronal loss between 4m and 10m in the 5XFAD
mice, showing that all significant loss in the subiculum occurs by 4m of age in this mouse
model.
• Overall Nurr1 protein and transcript levels are decreased in the hippocampi of 5XFAD
mice compared to the non-transgenic control by 4 months of age (Fig 3). This indicates
that changes in Nurr1 expression in the 5XFAD are following a similar time course to
neuronal death in this model.
• While it is known that Nurr1 can play anti-inflammatory roles in microglia and astrocytes
(Saijo et al. 2009), we find that Nurr1 is mainly expressed in neurons in the subiculum of
both wild-type and transgenic mice (Fig 4). We find it likely that Nurr1 is playing a
neuronal role during the development of AD pathology in these mice.
Figure 2. Neuronal death
occurs in the subiculum by 4M
in 5XFAD mice. Neuron counts in
4M and 10M subiculum of WT and
transgenic mice. Neurons were
immunolabeled with NeuN and
quantified (n=4-6/group). The
number of neurons significantly
decreased in the 4M transgenic
subiculum compared to WT of the
same age. There was no significant
neuronal loss between 4M transgenic
and 10M transgenic mice (*p≤0.05).
Figure 1. 5XFAD mice experience microgliosis and increases in plaque
area during aging. Immunostaining of plaques and microglia in the subiculum of
4M (a) and 10M (b) mice. Amyloid-β plaques and microglia are increased in the
subiculum of transgenic mice (10X magnification). (c) Microglial area analysis of 4M
and 10M WT and 5XFAD subiculum labeled using Iba1 and analyzed using ImagePro
Plus (n=4-6/group). Percent microglial area increased from WT to transgenic but did
not significantly change with age (*p≤0.05) (d) Senile plaque area in 4M and 10M
5XFAD subiculum labeled with 6e10 and analyzed using ImagePro Plus (n=4-6/group).
Plaque area trend increases with brain age (p=.07).
(a) 4M B65XFAD
(b) 10M B65XFAD
Iba1
Iba1
6e10
6e10
Merged
Merged
(c) (d)
Figure 3. Nurr1 transcripts and protein decrease in the hippocampus
in 5XFAD mice. (a) qPCR of RNA from 4M and 10M animals including 5XFAD and
non-transgenic homogenized hippocampus (n=4-6). (b) Analyzed Nurr1 western of 4M
and 10M WT and 5XFAD protein from hippocampal homogenates (n=4-6).
Figure 4. Nurr1 colocalizes with neurons in 4M hippocampus.
Immunolabeling in the subiculum with Nurr1 and NeuN along with a DAPI counterstain.
Includes 4M transgenic (a) and WT (b) mice (10X magnification).
Nurr1
Nurr1
NeuN
NeuN
Merged
DAPI
Merged
DAPI
(a) B65XFAD
(b) WT
(b)(a)