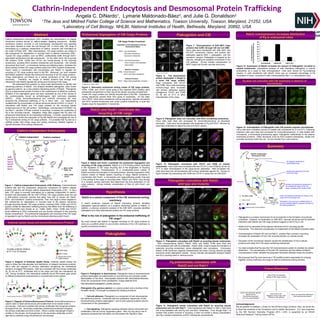
1. Clathrin-Independent Endocytosis and Desmosomal Protein Trafficking
Angela C. DiNardo1
, Lymarie Maldonado-Báez2
, and Julie G. Donaldson2
1
The Jess and Mildred Fisher College of Science and Mathematics, Towson University, Towson, Maryland, 21252, USA
2
Laboratory of Cell Biology, NHLBI, National Institutes of Health, Bethesda, Maryland, 20892, USA
Plakoglobin and CIE
Pg preferentially colocalizes with
Rab22 and not Rab11
Abstract
Clathrin-independent Endocytosis
Antibody Uptake Assay and Immunoflourescence Protocol
Clathrin-independent endocytosis (CIE) mediates the internalization of integral
membrane proteins devoid of clathrin-adaptor targeting sequences. Multiple
membrane proteins involved in cell homeostasis, development and signaling
have been reported to enter the cell through CIE. In HeLa cells, CIE cargo is
internalized by a pathway independent of clathrin, dynamin and modulated by
the small GTPase Arf6. After internalization, CIE-cargo proteins are sorted to
late endosomes for degradation or recycled back to plasma membrane (PM).
Typical CIE cargo proteins, such as MHCI, preferentially traffic through EEA1-
associated endosomes and late endosomes. On the other hand, the long-lived
PM proteins CD44, CD98, and CD147 are routed directly to the recycling
endosomes, avoiding EEA1-positive endosomes and lysosomes. We recently
discovered that Hook1, a microtubule binding and tethering factor, mediates the
direct sorting of these CIE cargo proteins away from EEA1-positive endosomes.
Hook1 works in coordination with microtubules and Rab22 to regulate the
recycling of CIE cargo proteins back to the PM. Moreover, our work revealed
that Rab22 depletion impairs the sorting and recycling of all CIE cargo proteins.
These observations put Rab22 as a central component of the CIE sorting
machinery. Therefore, we sought to identify proteins that through their
interactions with Rab22 regulate the sorting and recycling of CIE cargo.
We addressed this question by using mass spectrometry to identify new
interacting partners of Rab22. This analysis identified plakoglobin, (also known
as gamma-catenin), as a new putative interacting partner of Rab22. Plakoglobin
(Pg) is a desmosomal protein involved in the maintenance of adherins junctions.
Mutation of the plakoglobin gene has been implicated as one of the causes of
the cardiomyopathy known as arrhythmogenic right ventricular dysplasia
(ARDV). Using immunofluorescence and confocal microscopy we have
analyzed the endosomal trafficking of Pg in HeLa cells. Our experiments
revealed that Pg concentrates in internal structures devoid of EEA1 or Lamp1. It
colocalizes with Rab22 and CIE cargo proteins on the recycling tubular
endosomes and on the cell surface. Interestingly, overexpression of Rab22, but
not Rab11, (another GTPase involved in recycling), increases the localization of
Pg in the recycling tubules. Altogether, these data suggest that Pg uses CIE
endosomal membranes for its intracellular trafficking. Currently, experiments are
being done to confirm the interaction of Pg with Rab22 and investigate the role of
Pg in endosomal sorting of CIE cargo proteins. It is possible that Pg acts as a
scaffold protein that brings cargo and sorting machinery (Rab22) together in
specific domains of the sorting endosomes.
EEA1
ERC
LE
Clathrin-independent Clathrin-mediated
Arf6Q67L
MHCI, CD59
CD55, Tac
CD44, CD98
CD147
Rab5
Rab35Arf6
Arf6
Rab35
Rab22
Rab10
Rab8
Rab11 Rab22
Hook1
SE
Tfn
Figure 1. Clathrin-independent Endocytosis (CIE) Pathway. Transmembrane
proteins that lack the cytoplasmic sequences recognized by clathrin adaptor
proteins enter the cell through clathrin-independent endocytosis (CIE). In HeLa
cells, CIE cargo is normally internalized by a pathway independent of clathrin,
dynamin and associated with the Arf6 GTPase. CIE cargo proteins enter the cell
in Arf6-positive endocytic vesicles that either fuse with or mature into Rab5-,
EEA1- and transferrin- positive endosomes. Then, the cargo is either targeted to
late endosomes for degradation or recycled back to the plasma membrane
through recycling tubules. CIE cargo proteins, including CD44, CD98, and CD147
(green), exhibit an alternative trafficking itinerary that differs from the trafficking of
MHCI/Tac/CD59 (red) (Eyster et. al., Traffic, 2009). CD44, CD98, and CD147
avoid trafficking to EEA1-endosomes and are directly delivered to the recycling
tubular compartment. The endosomal segregation and recycling of the CIE cargo
is regulated in part by Rab22 and the microtubule tethering protein Hook1.
Figure 2. Diagram of Antibody Uptake Assay. Antibody uptake assays are
used to follow the internalization and distribution of integral membrane proteins.
HeLa cells are exposed to primary antibodies recognizing the extracellular
domains of integral PM proteins. Cells are incubated with the primary antibodies
for 30 min at 37°C. Antibodies bind to the cargo and both are internalized via
CIE. During incubation, the internalized antibody-bound PM protein gets sorted
to different endosomal compartments or recycled back to the surface.
Surface antigens
Primary antibody introduced
Primary antibody binds
to specific antigen
Seconday antibodies
recognizes and bind to
primary
Fluorophores fluoresce
under light emitted by
microscope
Endosomal Segregation of CIE Cargo Proteins
Regulation of Cell Interactions with the
Extracellular Matrix
CD44 – cell-cell adhesion
cell-matrix interactions
hyaluronan metabolism
CD147 – cell-ECM interactions
tumor invasion and metastasis
inflammation
Nutrient Transport
CD98 - nutrient transport
role in cell matrix interactions
Immune Function
MHCI – Major Histocompatibility Complex I
Eyster et al. Traffic, 2009
CIE Cargo Protein Functions
Figure 4. Alternative endosomal sorting routes of CIE cargo proteins.
CD44, CD98, and CD147 avoid going to the classical EEA1-positive early
endosomal compartment on their way to the tubular recycling compartment.
These CIE cargo proteins are directly recycled back to the PM. Cytoplasmic
sorting sequences prevent them from going to lysosomes. On the other
hand, MHCI, which lacks cytoplasmic sorting sequences, typically colocalizes
with EEA1-positive endosomes and Lamp1-positive endosomes, a route that
targets cargo for degradation in lysosomes.
Rab22 and Hook1 coordinate the
recycling of CIE cargo
Figure 5. Rab22 and Hook1 coordinate the endosomal segregation and
recycling of CIE cargo proteins. Rab22 is a GTP-binding protein implicated
in the regulation of CIE cargo recycling and in the biogenesis of recycling
tubular endosomes. Overexpression of a constitutively-active mutant of
Rab22 promotes the formation of recycling tubules, whereas expression of the
inactive mutant of Rab22 impairs recycling of cargo. Rab22 functions in
coordination with Hook1, a microtubule and cargo-tethering protein that aids
in the sorting of the cargo to recycling tubes. Rab22 and Hook1 may be part
of a complex sorting machinery that determines the final destination of CIE
cargo proteins. Arrows indicate colocalization of Rab 22 with Hook1 and
CD147.
Rab22 is a central component of the CIE endosomal sorting
machinery.
A recent proteomic analysis of Rab22 interacting proteins identified
plakoglobin (gamma-catenin) as a putative interacting partner of Rab22. In
addition, a previous proteomic analysis of the Arf6 Q67L vacuoles identified
plakoglobin and other desmosomal proteins as CIE cargo proteins.
What is the role of plakoglobin in the endsomal trafficking of
CIE cargo?
Pg could interact with Rab22 to regulate recycling of CIE cargo proteins or
serve as a scaffold protein to recruit other members of the CIE machinery to
specific endosomal domains.
Hypothesis
Plakoglobin (Pg, gamma-catenin) is a cytsolic protein and a member of the
armadillo family. It is thought to possess the following functions:
1. Cell-cell adhesion: Plakoglobin is a component of both desmosomes
and adherins junctions. It interacts with the cytoplasmic sequences of both
transmembrane proteins (desmoglein 1 and 2) and cytosolic proteins (bind to
components of the cytoskeleton)
2. Cell-signaling: Plakoglobin signaling activities reduce cell growth and
proliferation (role as a tumor suppressor gene). Also, Pg may play a role in
apoptosis (programmed cell death) and decreased cell migration rates.
http://www.eheart.org/pages/01_cardiac_structure
CD147
CD98
Dsg2
Dsg2
Merge
Merge
Figure 7. Overexpression of Arf6 Q67L traps
proteins that traffic through CIE but not CME.
HeLa cells were transfected with Arf6 Q67L and
PH-GFP (membrane marker) and probed with
antibodies against MHC I, plakoglobin, and the
transferrin receptor. Plakoglobin is trapped by the
vacuole, indicating its possible involvement in the
CIE pathway. Arrows indicate colocalization of
MHCI cargo and Pg with vacuolar structures.
Figure 8. The desmosomal
protein desmoglein 2 (Dsg2) is
a novel CIE cargo protein.
Dsg2 colocalizes with CD147
and CD98. HeLa cells expressing
mCherry-Dsg2 were incubated
with primary antibodies against
endogenous CD147 and CD98
for 30 min at 37°C to allow
internalization of the cargo.
Figure 9. Plakoglobin does not colocalize with EEA1-containing endosomes.
HeLa cells were fixed and processed for immunofluorescence as described
previously. Cells were immunolabeled with antibodies to Pg and EEA1, followed by
secondary antibodies to detect Pg (red) and EEA1 (green).
MergePlakoglobin EEA1
Figure 10. Plakoglobin colocalizes with CD147 and CD98 on tubular
endosomes. HeLa cells were incubated with anti-CD147 or anti-CD98 for 30 min at
37°C to allow internalization of the cargo-bound antibodies. After incubation the
cells were fixed and immunostained with primary antibodies against Pg. Arrows on
figure indicate Pg colocalizing with CD98 and CD147 in tubes and not with EEA1.
EEA1
CD147CD98
Merge
Figure 11. Plakoglobin colocalizes with Rab22 on recycling tubular endosomes.
Cells overexpressing Rab22, Rab22 Q64L and Rab22 S19N were fixed and
immunolabeled with anti-Pg antibodies. Rab22 Q64L mutant causes the formation of
extensive recycling endosomes, where plakoglobin also colocalizes. Conversely,
Rab22 S19N (dominant negative) disassembles the tubes, forming internal punctate
structures where plakoglobin resides. Arrows indicate colocalization between Rab22
and Pg in reycling tubes or internal puncta.
Figure 12. Plakoglobin mostly colocalizes with Rab22 on recycling tubular
endosomes and not with Rab11. Cells transfected with Rab22 or Rab11 were fixed
and immunolabeled with anti-Pg and anti-EEA1 antibodies. Even though Rab11 is
another Rab protein involved in recycling, it does not exhibit a strong colocalization
with Pg in tubular endosomes or EEA1 endosomal compartments.
Plakoglobin EEA1 MergeGFP-Rab22
Plakoglobin EEA1 MergeGFP-Rab11
Figure 13. Expression of Rab22 increases the amount of Plakoglobin recruited to
tubular endosomes. Quantitative analysis of cells displaying plakoglobin in tubular
endosomes as a result of Rab overexpression. Cells were scored for Pg-containing
tubules. In cells transfected with Rab22, there was an increased percentage of Pg
localization to tubes, compared to cells overexpressing Rab11 or untransfected cells.
ControlNZ
Plakoglobin EEA1 Merge
Figure 14. Colocalization of Plakoglobin with CIE proteins requires microtubules.
HeLa cells were untreated (control) or treated with nocodazole for 2 h at 37°C. Following
treatment cells were fixed and processed for immunofluorescence. In cells treated with
nocodazole, a microtubule polymerizing agent, Plakoglobin redistributed to the plasma
membrane junctions. Other structures, such as EEA1-positive endosomes, cluster into
punctate structures within the cell. Arrows indicate Pg in cell junctions.
• Plakoglobin is a protein most known for its involvement in the formation of junctional
complexes. However, its localization on Arf6 Q67L vacuolar structures and its potential
interaction with Rab22 and CIE cargo proteins suggest a role for Pg in CIE.
• Rab22 and Pg colocalize at the plasma membrane and in recycling tubular
endosomes. The observed colocalization is independent of the Rab22 activation state.
• Overexpression of Rab22 WT and not Rab11, another Rab involved in recycling,
increases the localization of Pg in the recycling tubules vs. internal puncta.
• Disruption of the microtubule network causes the redistribution of Pg to cellular
junctions and away from CIE cargo-containing endosomes.
• Our data indicates that Pg uses the CIE endomembrane system to maintain its cellular
distribution. The colocalization of Pg with CIE machinery and the effects of Rab22
overexpression further validates the proposed interaction between these two proteins.
• We proposed that Pg could serve as a CIE scaffold protein responsible for bringing
together sorting machinery and cargo to distinct endosomal sorting domains.
No apparent specificity at
internalization step.
Sorting step determines the fate
of the CIE cargo.
CD44, CD147
Tac, MHCI, CD59
Rab5
EEA1
Clathrin-independent
Endocytosis (CIE)
LE
ERC
Rab5
Arf6
Sorting by
Acidic
Residues
Sorting Proteins (Hook1/Rab22)
Sorting event guided by sequences in
the cytoplasmic region of the CIE
cargo, including acidic cluster motifs.
Degradation
Recycling
Hook1 and Rab22 coordinate the sorting of CIE
cargo into the recycling route.
Plakoglobin may serve as a scaffold protein to
recruit or stabilize Rab22 on specific domains of
the sorting endosome.
Plakoglobin
Role of Pg in Endosomal
Sorting of CIE cargo proteins
Confirm the interaction between
Rab22 and Pg
•Biochemical Approach
•Co-Immunnoprecipitation
Role of Pg in CIE
•Depletion of Pg in HeLa cells
•Effects on CIE pathway
Study the endosomal trafficking of
other desmosomal proteins
Incubate antibody dilutions
at 37°C for 30 minutes
CD98 cargo
MHCI cargo
Mouse anti-human
CD98 antibody
Figure 3. Diagram of Immunoflourescence Protocol. Immunoflourescence is
used to stain or label cell structures and enable them to be viewed under a
microscope. Cells are incubated with primary antibodies that recognize proteins
throughout the cell. Secondary antibodies containing fluorophores recognize
the primary antibodies and bind to them. When a certain wavelenght of light is
emitted on the sample, the fluorophores on the secondary antibodies re-emit
light that is collected using a fluorescence microsocope.
Figure 6. Plakoglobin in desmosomes. Plakoglobin binds to transmembrane
proteins desmoglein and desmocollin on one terminus and cytosolic protein
desmoplakin on the other. Desmoplakin interacts with intermediate filaments,
which are components of the cytoskeleton. Image obtained from:
Plakoglobin
Plakoglobin colocalizes with Rab22
wild type and mutants
GFP-Rab22
Plakoglobin EEA1 Merge
GFP-Rab22S19NGFP-Rab22Q64L
Pg does not colocalize with CIE machinery in absence of
microtubules
Summary
Rab22 overexpression increases distribution
of Pg to endosomal tubes
Future Directions
Acknowledgements
We are grateful to Kathleen J Green for the mCherry-Dsg2 construct. Also, we would like
to thank the members of the Donaldson Lab for helpful discussions. A.D. was supported
by the NIH Summer Internship Program 2013. L.M.B. is supported by an NHLBI
Intramural Research Training Award (IRTA).
Hook1 MergeCD147
GFP-Rab22
GFP-Rab22(Q64L)GFP-Rab22(S19N)
Plakoglobin
Fraser et al. JCB, 2001