Wolf et al. "Graph abstraction reconciles clustering with trajectory inference through a topology preserving map of single cells"
1.
Wolf, F. Alexander,et al.
"Graph abstraction reconciles clustering with
trajectory inference through a topology preserving
map of single cells.”
bioRxiv preprint (2017).
報告者:鈴木良平
tmd-gpat Journal Club Apr.6, 2018



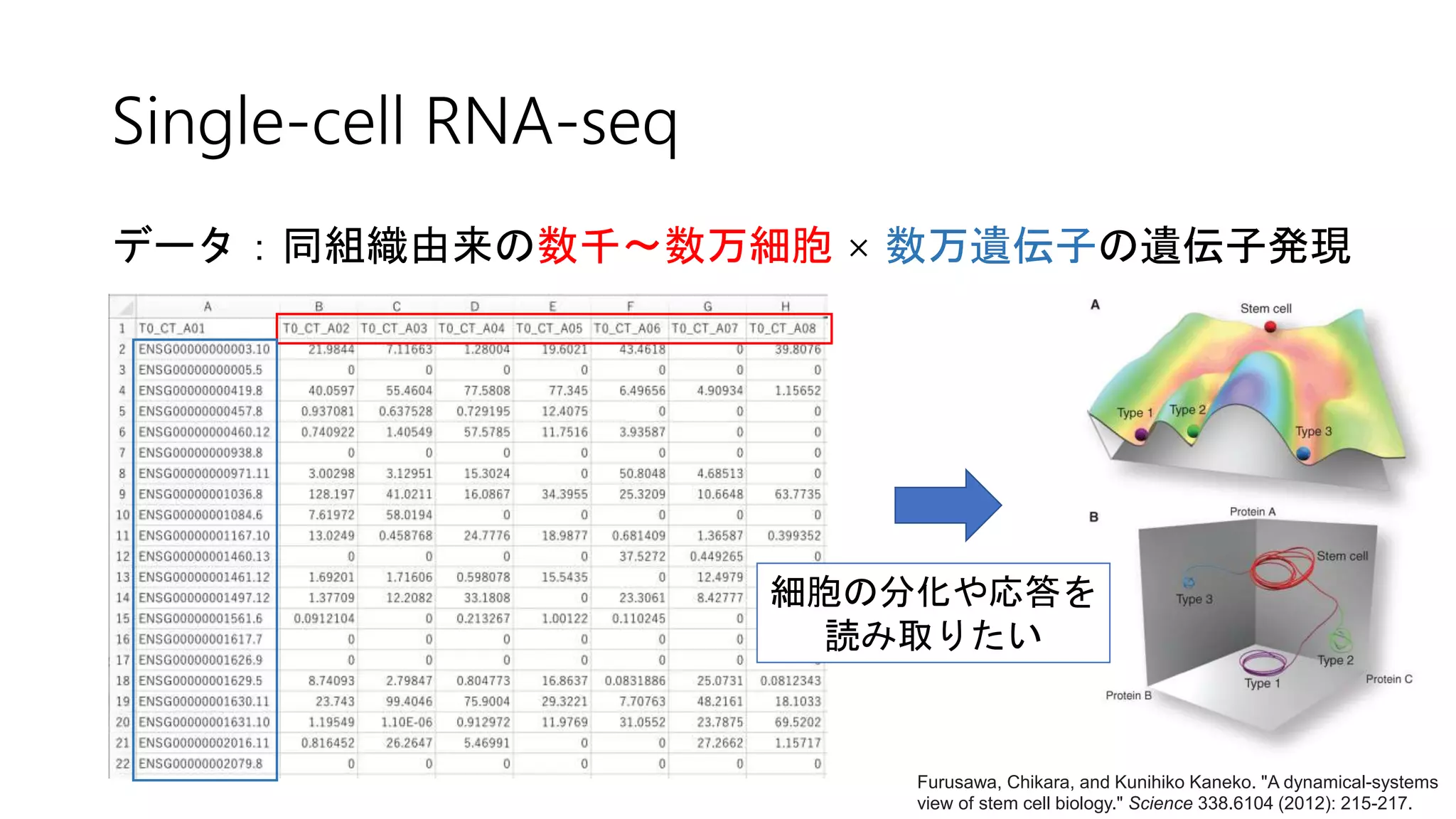

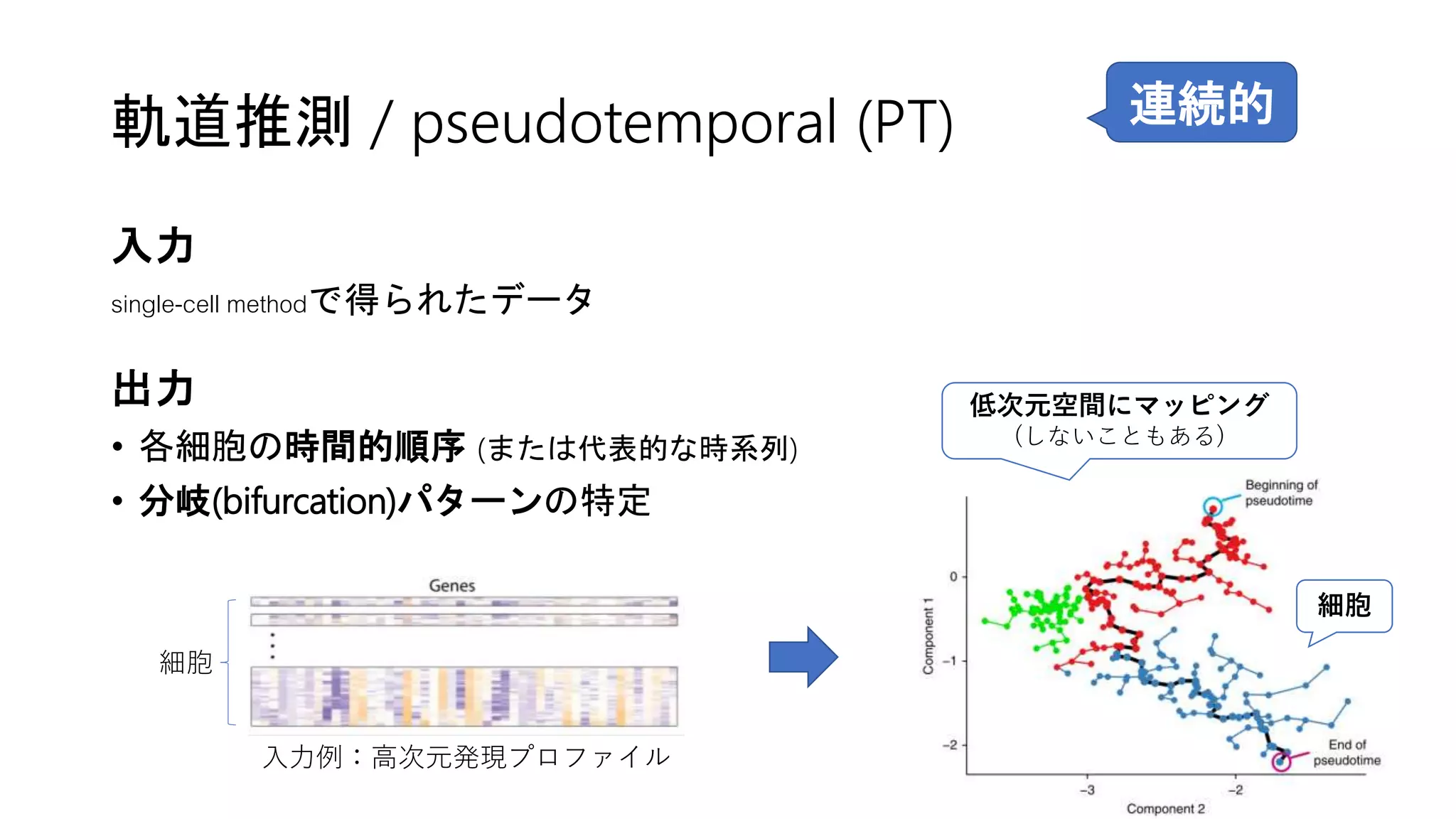
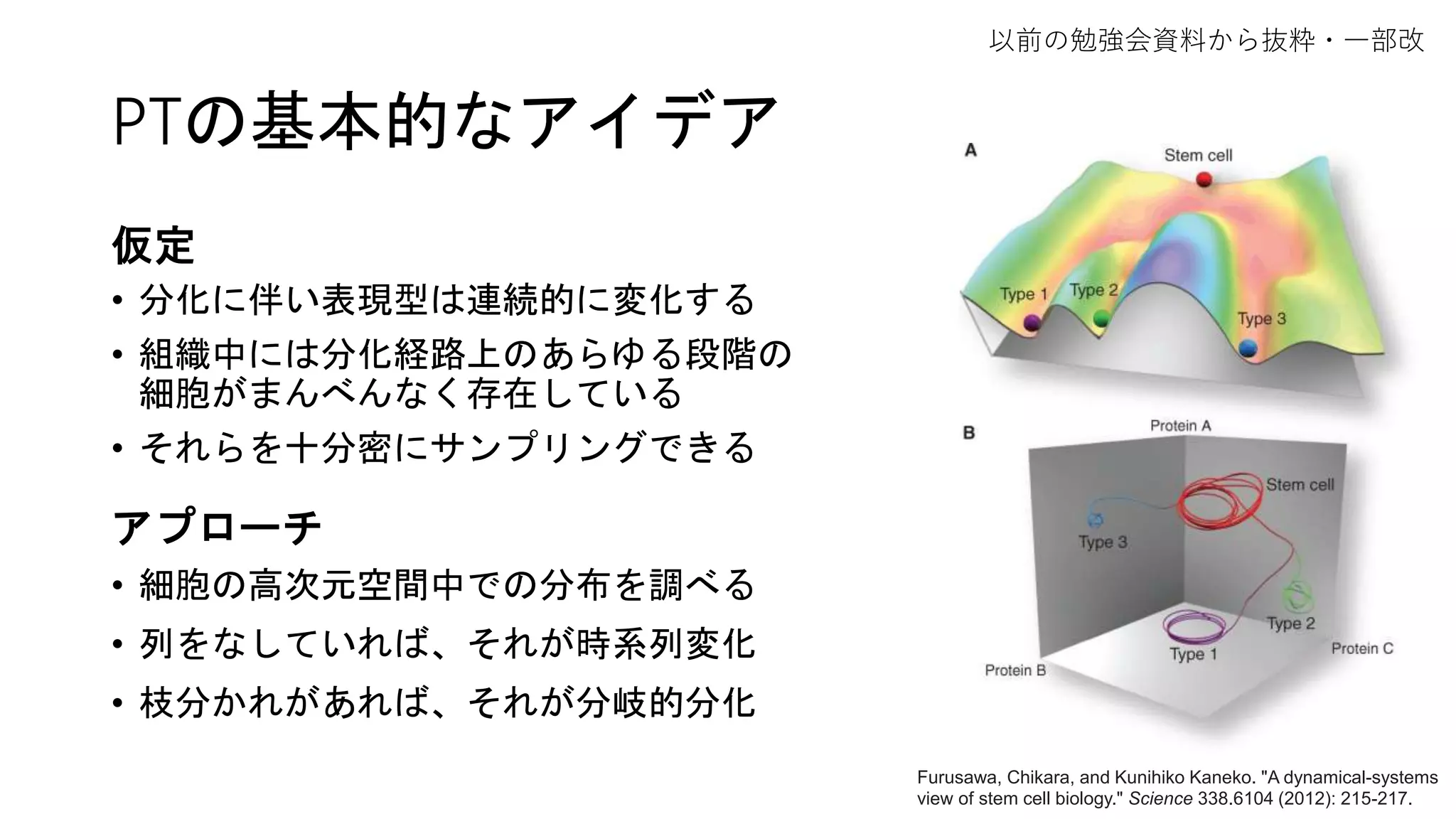

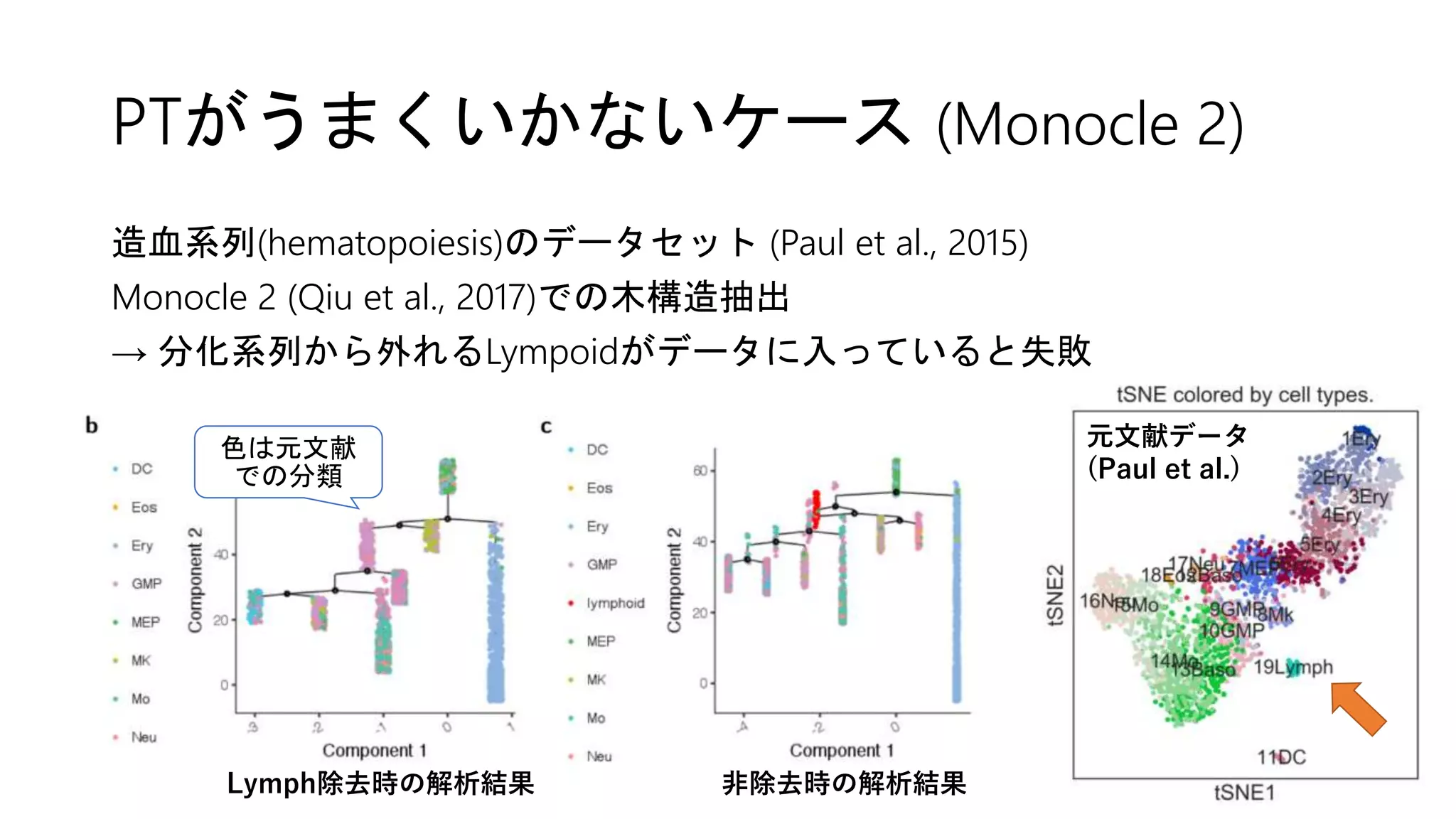

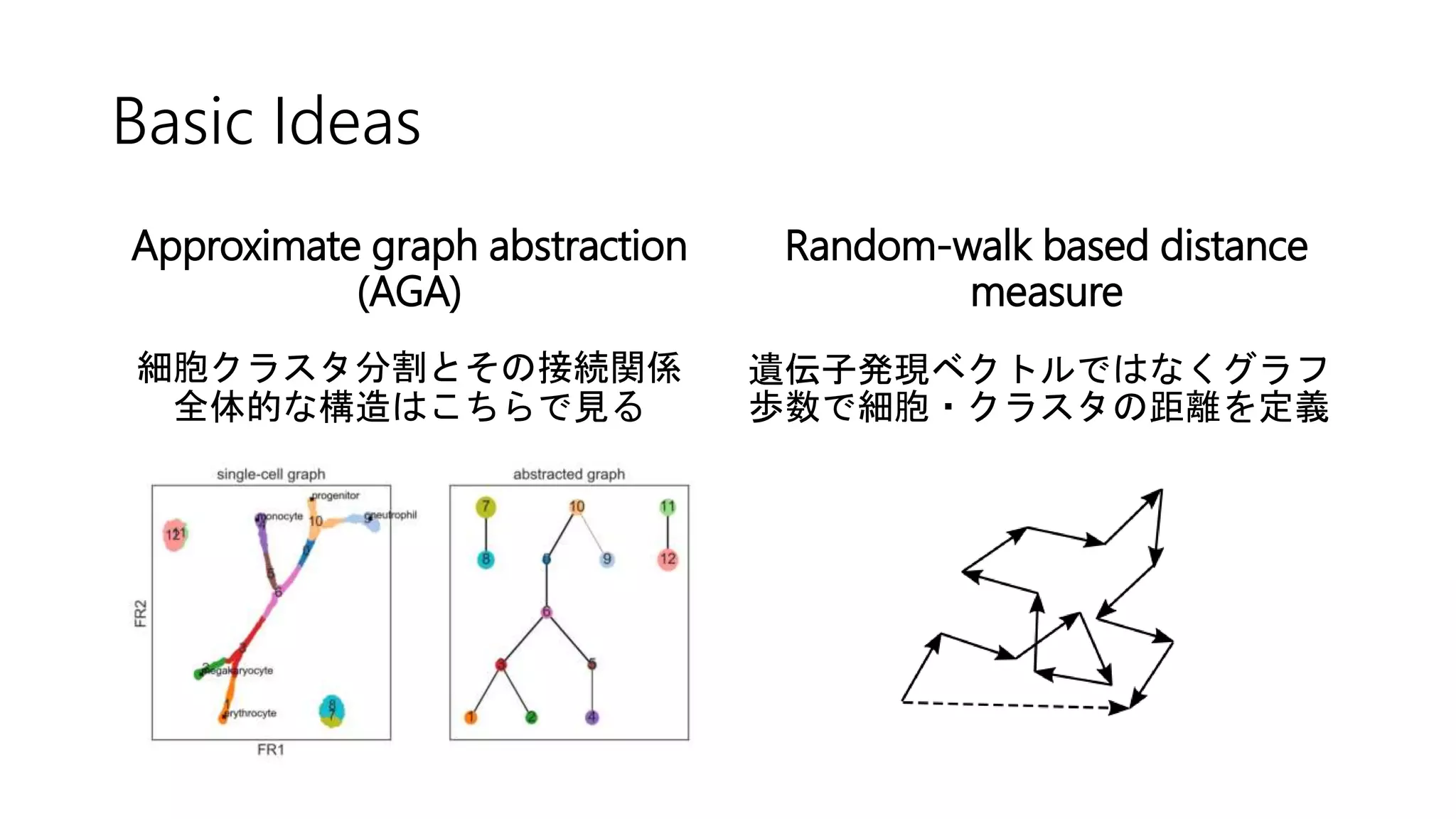
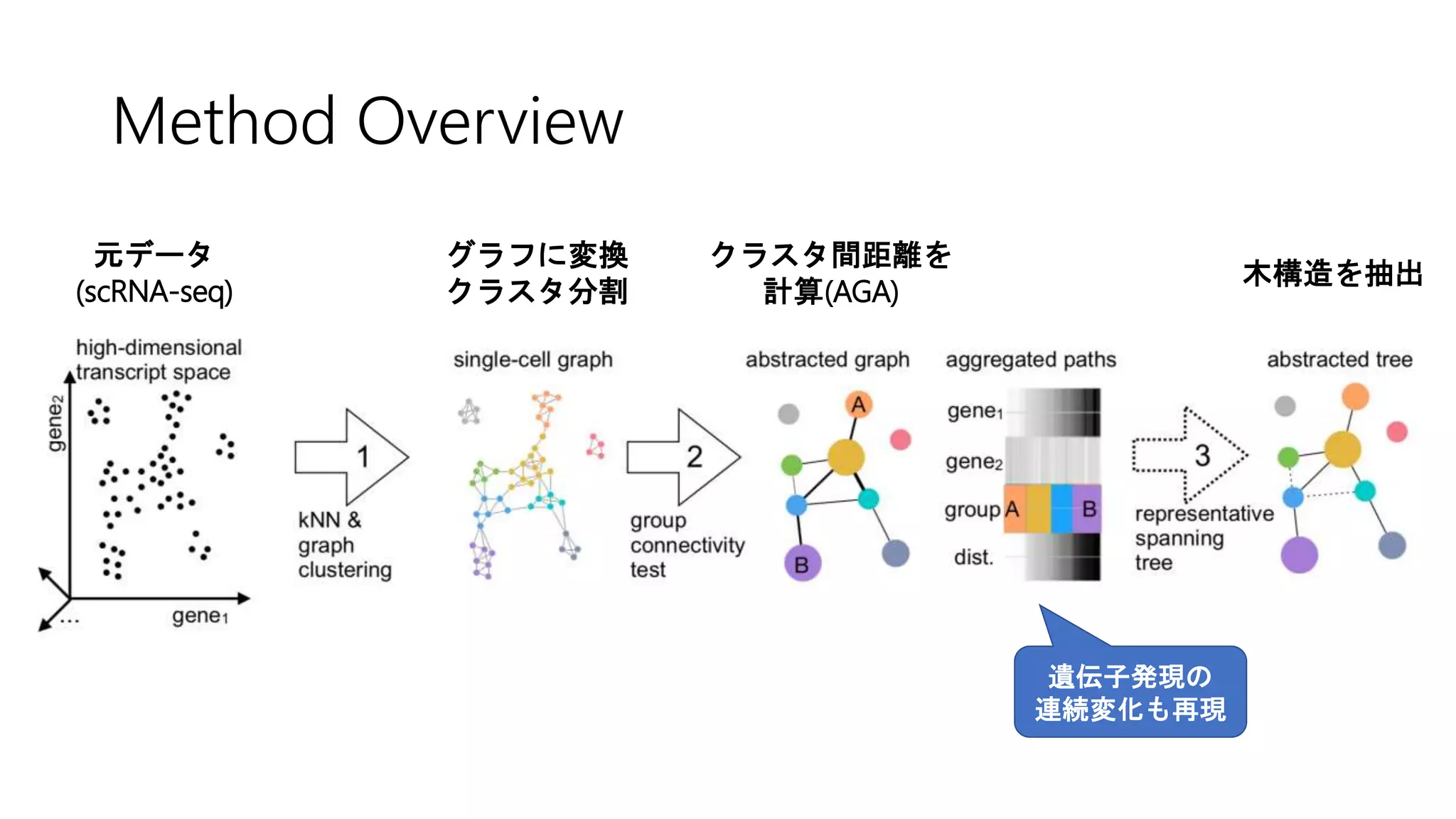

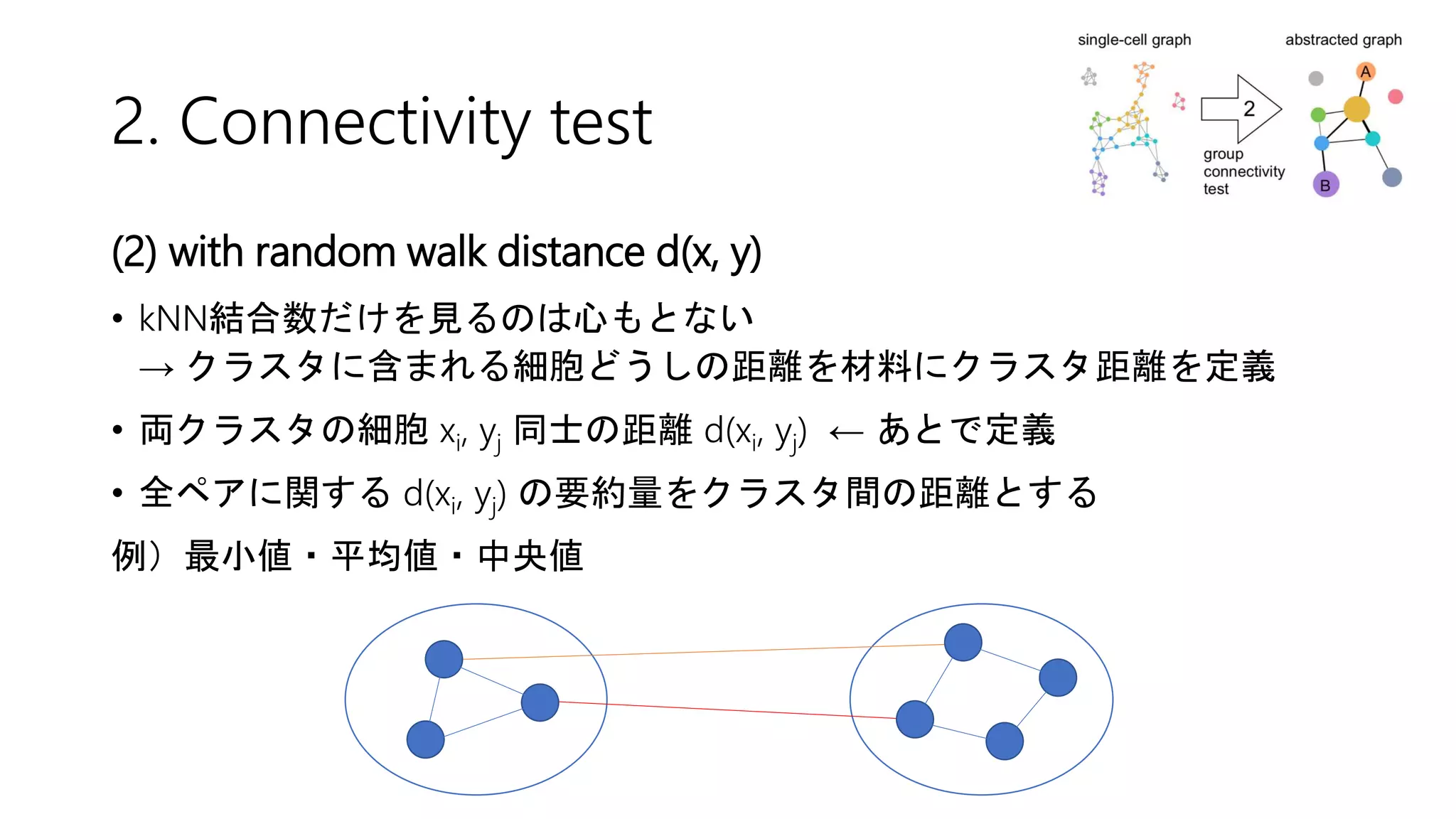
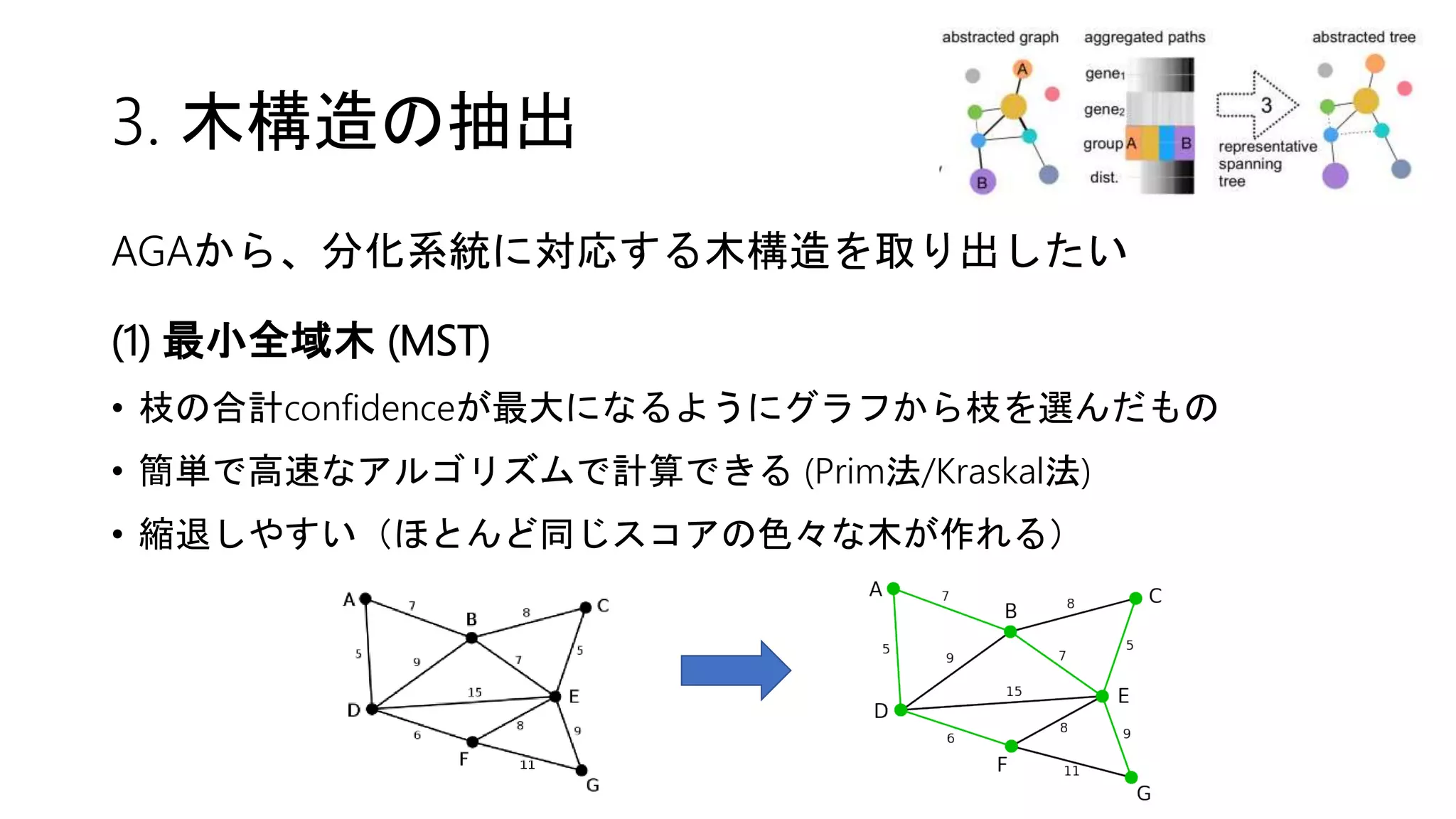
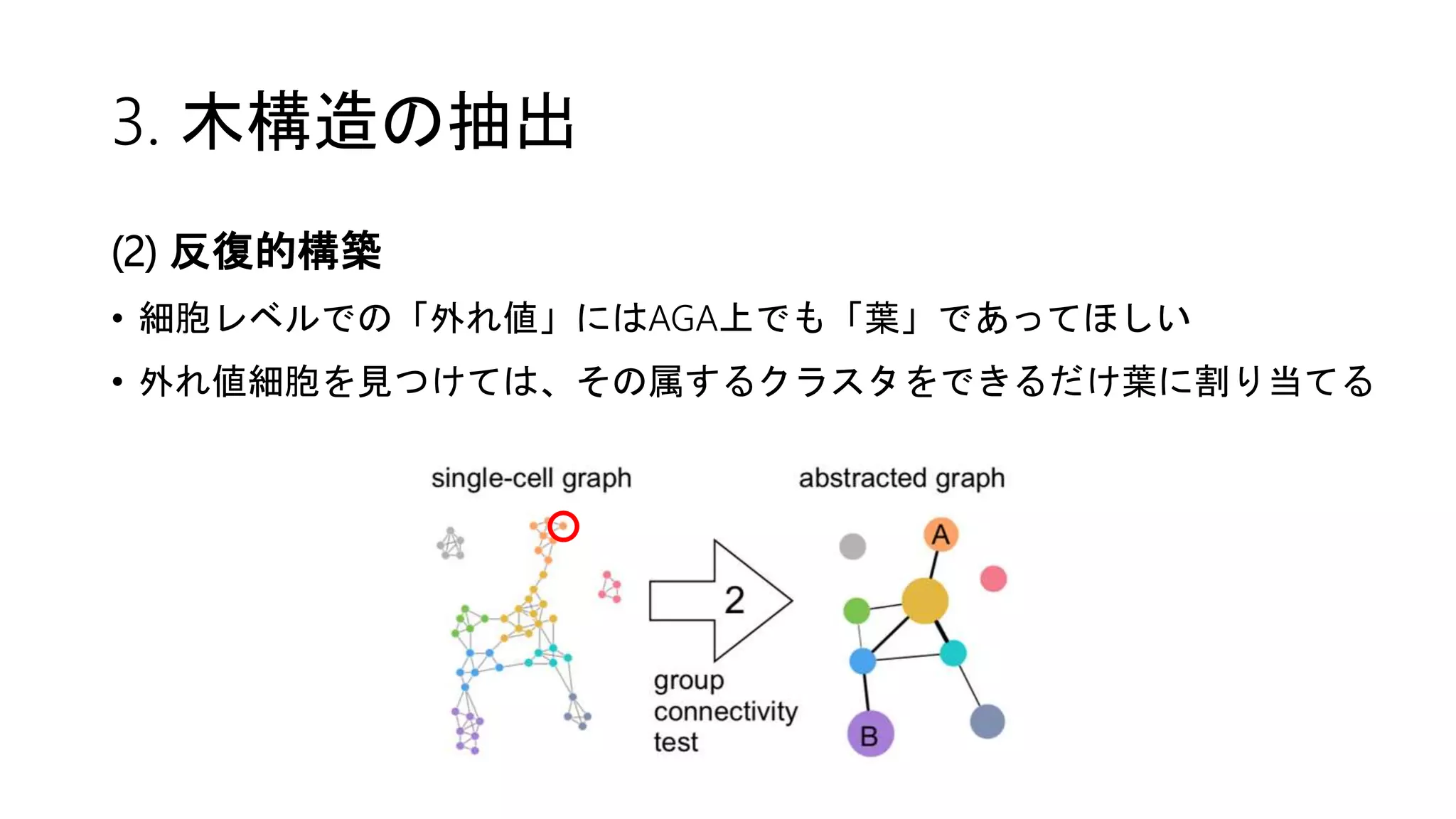
![2. Connectivity test
Partition同士の結合(↔距離)はどのように定義する?
(1) edge statistics
• partition i の関係する辺と全辺数との比 = edge frequency θi
• もしもpartition iとjが特に結合関係になければ、
相互接続数は期待値 E[kij] = Nθiθj のベルヌーイ分布に従う(粗い仮定)
→ 結合の確信度confidenceは二項検定で評価できる!
10%の辺が
集まった
partition
20%の辺が
集まった
partition
偶然だけなら
2%の辺が結ぶ](https://image.slidesharecdn.com/20180406suzuki-180406155409/75/Wolf-et-al-Graph-abstraction-reconciles-clustering-with-trajectory-inference-through-a-topology-preserving-map-of-single-cells-16-2048.jpg)


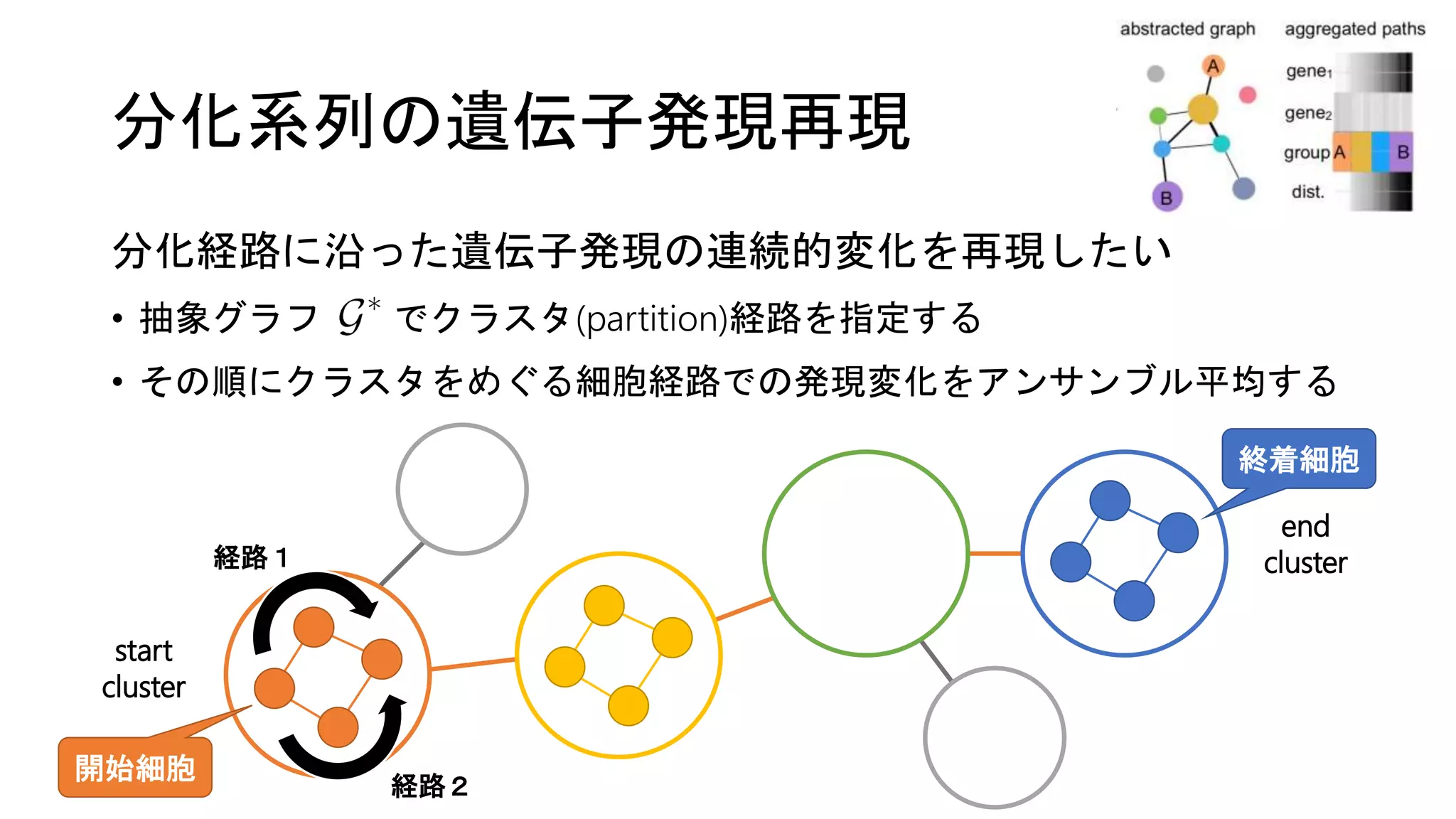

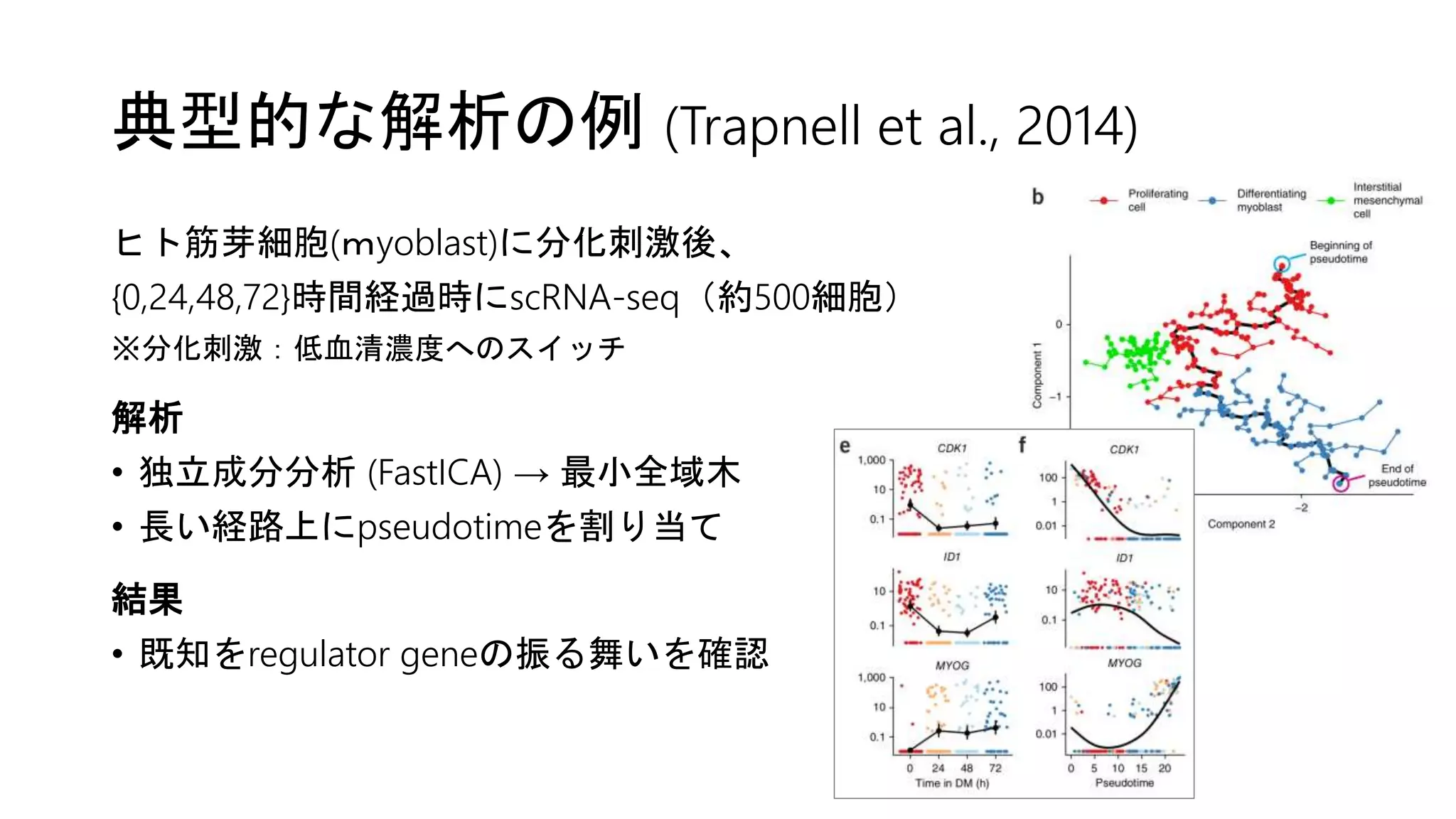
![実データでの評価
造血系列の3つのデータセットを使用
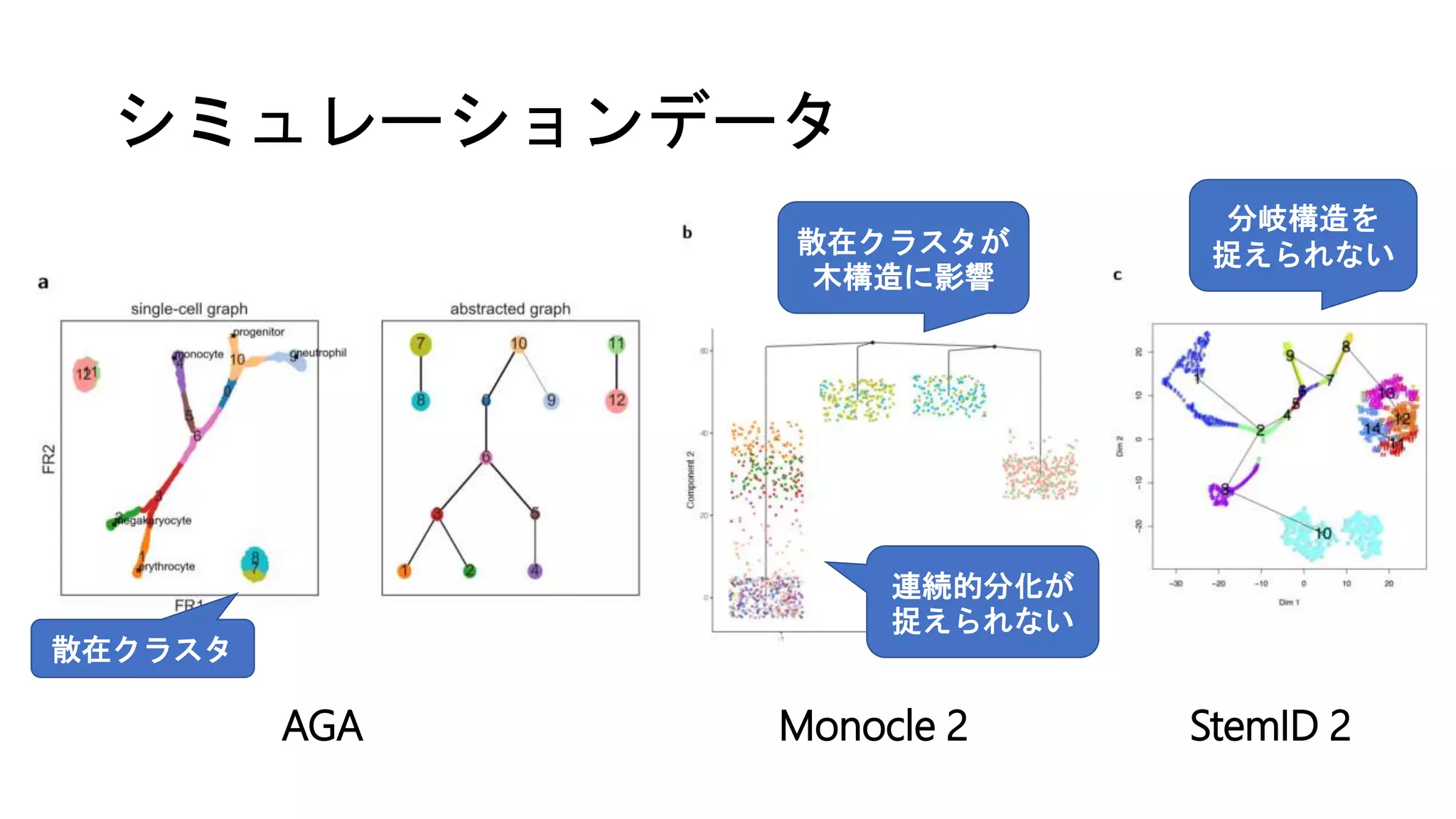
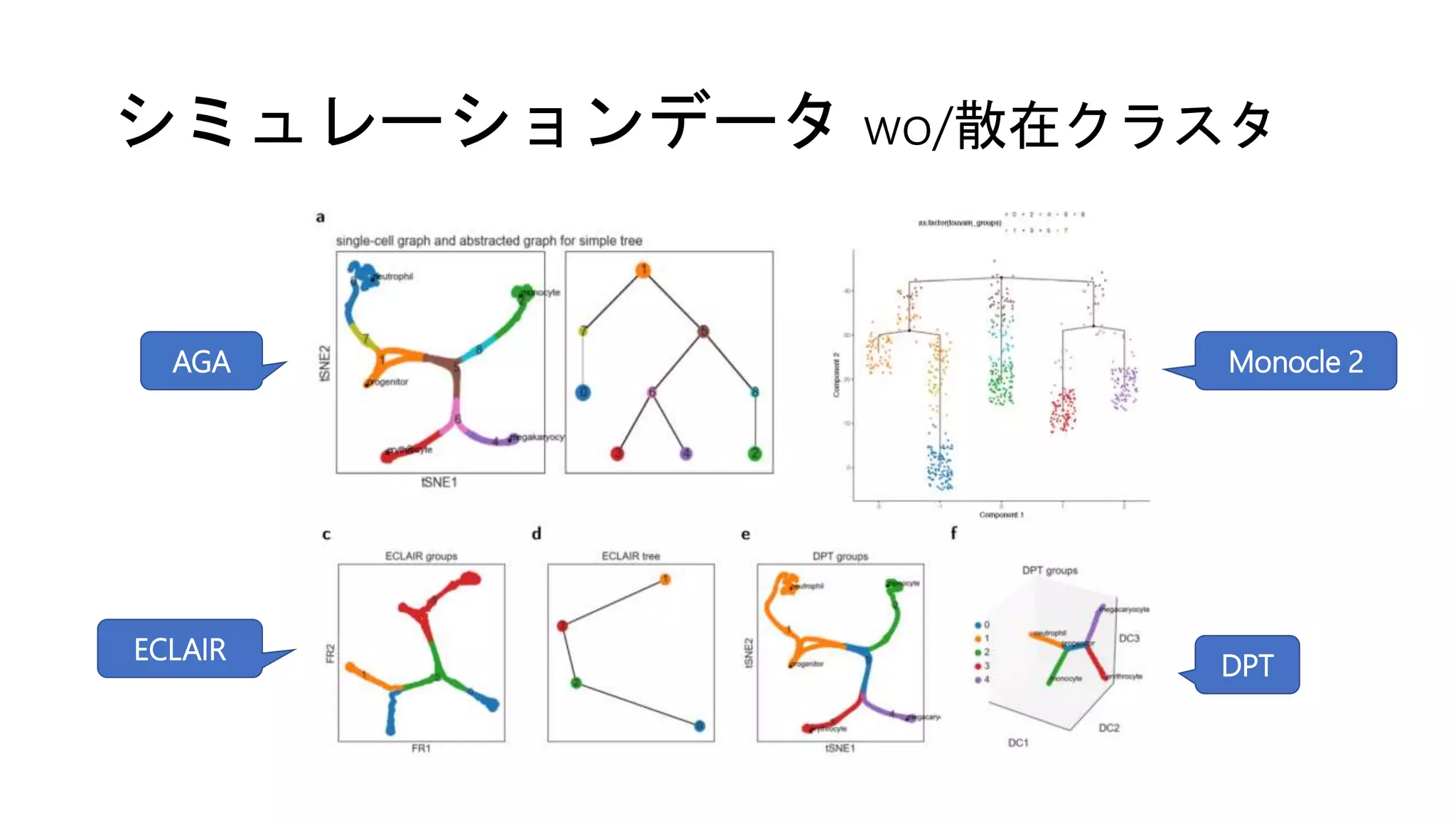
(1) シミュレーション [Moignard et al. ‘15]
• 11遺伝子のBoolean network (e.g., EKLF = Gata1 & not Fli1)
• ODEに変換 → 分化時の連続的な発現変化をシミュレーション
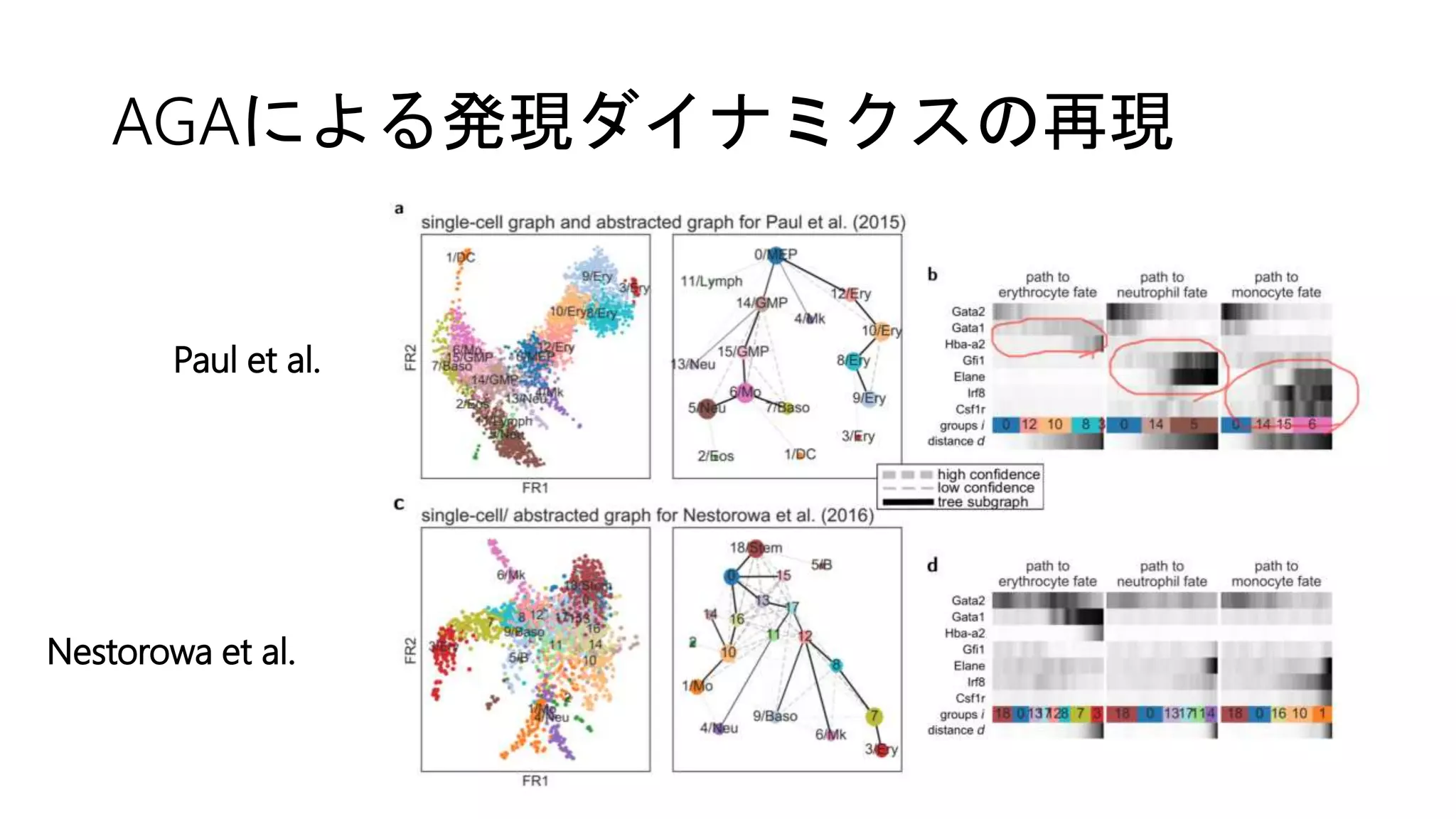
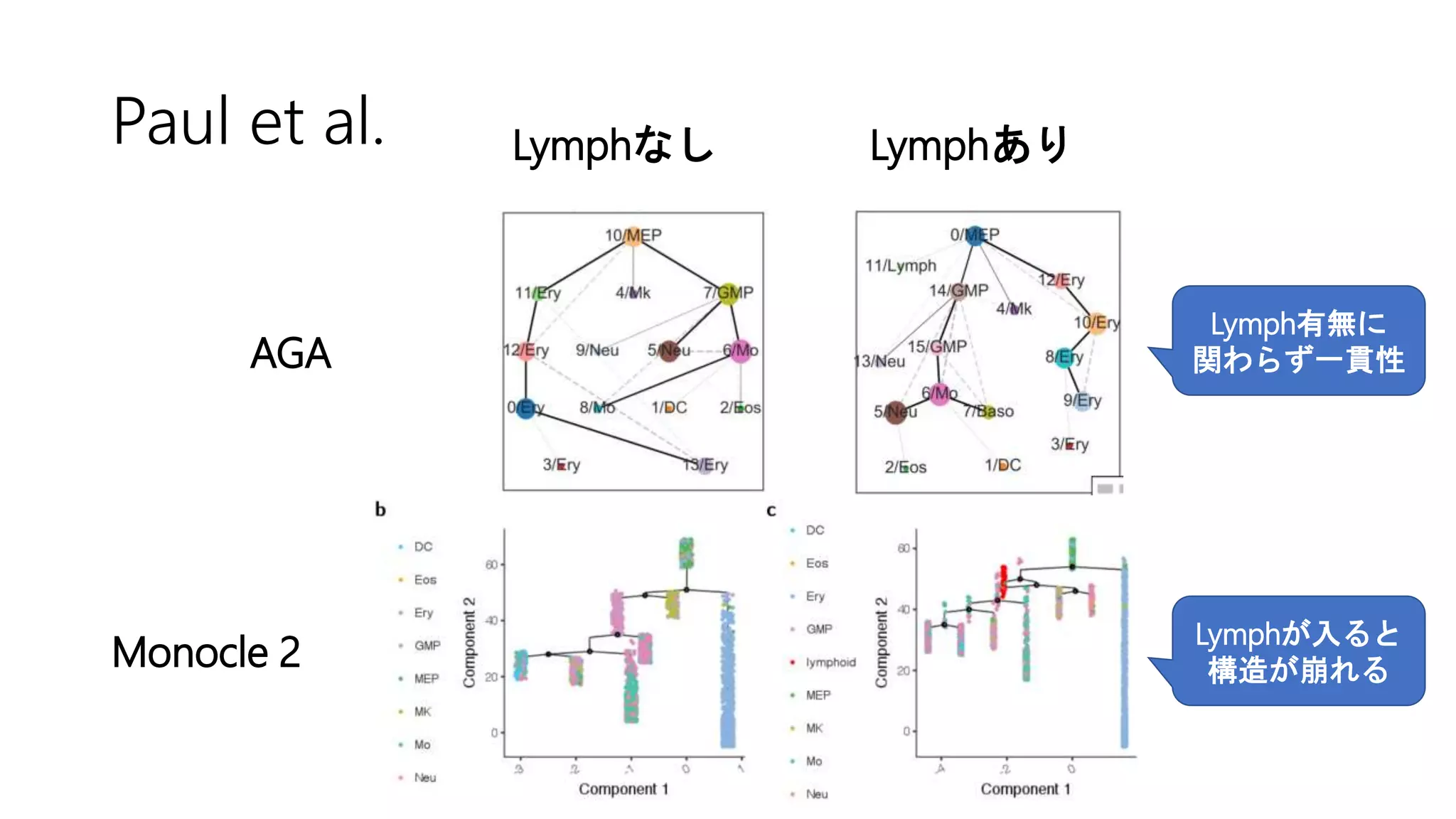
(2) Paul et al. 2015
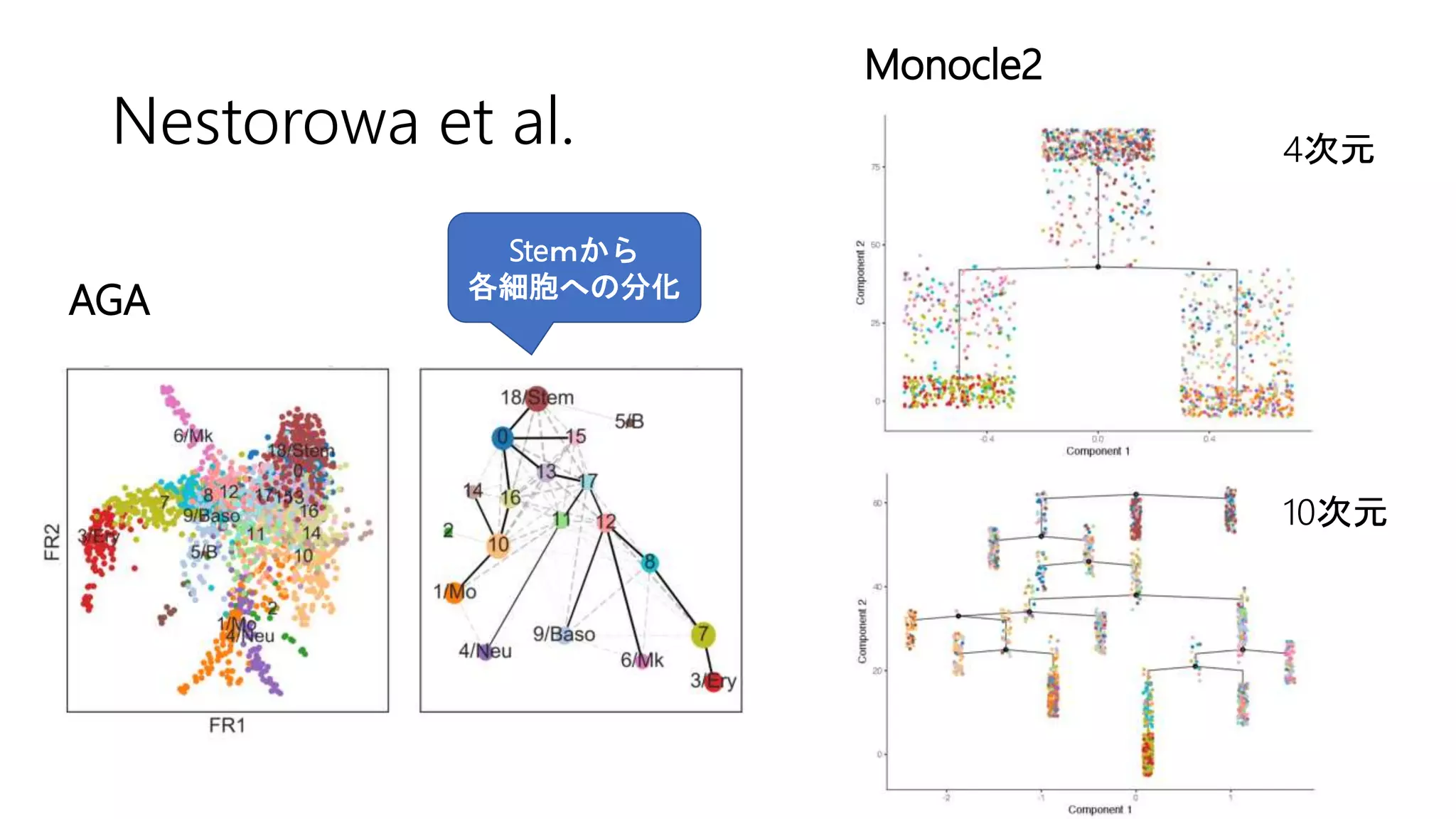
(3) Nestorowa et al. 2016
• マウスでのscRNA-seqデータセット](https://image.slidesharecdn.com/20180406suzuki-180406155409/75/Wolf-et-al-Graph-abstraction-reconciles-clustering-with-trajectory-inference-through-a-topology-preserving-map-of-single-cells-24-2048.jpg)