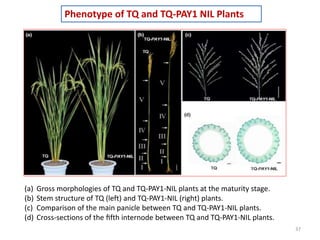
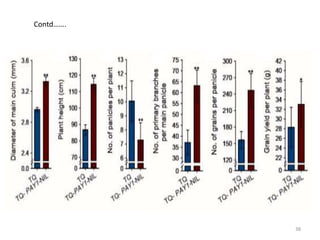
This document summarizes a study on the rice gene PAY1 and its effects on plant architecture. PAY1 was identified through mutagenesis of a wild rice introgression line. The PAY1 mutant showed changes to traits like plant height, tiller number, and panicle morphology. PAY1 was mapped and cloned, found to encode a protein that regulates polar auxin transport. Overexpression of PAY1 in the introgression line replicated the mutant phenotype. Introduction of PAY1 into an elite rice variety through backcrossing improved plant architecture and increased yield. The identification of PAY1 provides insights into molecular mechanisms of plant architecture and a tool for breeding higher-yielding rice varieties through modification of architecture.




























![32
3 groups of 5 days old Coleoptile Segments of 2cm length
incubation
Liq. Half strenght MS media, 100 rpm for 2hr
One end of apical/basal end is submerged in Half strenght MS media
To remove endogenous IAA
0.35% phytagel
500nm [3H]IAA
Dark
RT
2hr
NPA (N-1-naphthylphtalamic acid) applied to media for one group
Submerged end is washed with Half strenght MS media, 3times
Radioactivity of each section was counted by Liquid scintillation counter.
20 hr incubation in scintillation liquid
PAT Assay](https://image.slidesharecdn.com/a8e10229-ed54-40bc-b08d-4e06f31e13bc-160124081833/85/seminar-final-32-320.jpg)