Mathematical Foundations Of Neuroscience 1st Edition G Bard Ermentrout
Mathematical Foundations Of Neuroscience 1st Edition G Bard Ermentrout
Mathematical Foundations Of Neuroscience 1st Edition G Bard Ermentrout
Mathematical Foundations Of Neuroscience 1st Edition G Bard Ermentrout
Mathematical Foundations Of Neuroscience 1st Edition G Bard Ermentrout
1.
Mathematical Foundations OfNeuroscience 1st
Edition G Bard Ermentrout download
https://ebookbell.com/product/mathematical-foundations-of-
neuroscience-1st-edition-g-bard-ermentrout-2536774
Explore and download more ebooks at ebookbell.com
2.
Here are somerecommended products that we believe you will be
interested in. You can click the link to download.
Mathematical Foundations Of Data Science Using R 2nd Edition 2nd Frank
Emmertstreib
https://ebookbell.com/product/mathematical-foundations-of-data-
science-using-r-2nd-edition-2nd-frank-emmertstreib-46539954
Mathematical Foundations Of Data Science Using R Frank Emmertstreib
Salissou Moutari Matthias Dehmer
https://ebookbell.com/product/mathematical-foundations-of-data-
science-using-r-frank-emmertstreib-salissou-moutari-matthias-
dehmer-46548862
Mathematical Foundations Of Data Science Tomas Hrycej Berhard
Bermeitinger
https://ebookbell.com/product/mathematical-foundations-of-data-
science-tomas-hrycej-berhard-bermeitinger-48176422
Mathematical Foundations Of Software Engineering Gerard Oregan
https://ebookbell.com/product/mathematical-foundations-of-software-
engineering-gerard-oregan-49997796
3.
Mathematical Foundations OfData Science Using R Frank Emmertstreib
Salissou Moutari Matthias Dehmer
https://ebookbell.com/product/mathematical-foundations-of-data-
science-using-r-frank-emmertstreib-salissou-moutari-matthias-
dehmer-50347612
Mathematical Foundations Of Imaging Tomography And Wavefield Inversion
Anthony J Devaney
https://ebookbell.com/product/mathematical-foundations-of-imaging-
tomography-and-wavefield-inversion-anthony-j-devaney-50399852
Mathematical Foundations Of Quantum Statistics A Y Khinchin
https://ebookbell.com/product/mathematical-foundations-of-quantum-
statistics-a-y-khinchin-51211064
Mathematical Foundations Of Big Data Analytics 1st Ed 2021 Vladimir
Shikhman
https://ebookbell.com/product/mathematical-foundations-of-big-data-
analytics-1st-ed-2021-vladimir-shikhman-51589028
Mathematical Foundations Of Quantum Mechanics New Edition New John Von
Neumann Editor Nicholas A Wheeler Editor Robert T Beyer Editor
https://ebookbell.com/product/mathematical-foundations-of-quantum-
mechanics-new-edition-new-john-von-neumann-editor-nicholas-a-wheeler-
editor-robert-t-beyer-editor-51946636
6.
Interdisciplinary Applied Mathematics
Volume35
Editors
S.S. Antman J.E. Marsden
L. Sirovich S. Wiggins
Geophysics and Planetary Sciences
Imaging, Vision, and Graphics
D. Geman
Mathematical Biology
L. Glass, J.D. Murray
Mechanics and Materials
R.V. Kohn
Systems and Control
S.S. Sastry, P.S. Krishnaprasad
Problems in engineering, computational science, and the physical and biological sciences
are using increasingly sophisticated mathematical techniques. Thus, the bridge between
the mathematical sciences and other disciplines is heavily traveled. The correspond-
ingly increased dialog between the disciplines has led to the establishment of the series:
Interdisciplinary Applied Mathematics.
The purpose of this series is to meet the current and future needs for the interaction between
various science and technology areas on the one hand and mathematics on the other. This
is done, firstly, by encouraging the ways that mathematics may be applied in traditional
areas, as well as point towards new and innovative areas of applications; and, secondly, by
encouraging other scientific disciplines to engage in a dialog with mathematicians outlining
their problems to both access new methods and suggest innovative developments within
mathematics itself.
The series will consist of monographs and high-level texts from researchers working on
the interplay between mathematics and other fields of science and technology.
G. Bard Ermentrout David H. Terman
Mathematical Foundations
of Neuroscience
123
9.
G. Bard Ermentrout
Universityof Pittsburgh
Dept. Mathematics
301 Thackeray Hall
Pittsburgh, Pennsylvania 15260
USA
bard@pitt.edu
David H. Terman
Ohio State University
Dept. Mathematics
231 W. 18th Ave.
Columbus, Ohio 43210
USA
terman@math.ohio-state.edu
Editors
S.S. Antman
Department of Mathematics
and
Institute for Physical Science
and Technology
University of Maryland
College Park, MD 20742, USA
ssa@math.umd.edu
J.E. Marsden
Control and Dynamical Systems
Mail Code 107-81
California Institute of Technology
Pasadena, CA 91125, USA
marsden@cds.caltech.edu
L. Sirovich
Division of Applied Mathematics
Brown University
Providence, RI 02912, USA
chico@camelot.mssm.edu
S. Wiggins
School of Mathematics
University of Bristol
Bristol BS8 1TW, UK
s.wiggins@bris.ac.uk
ISSN 0939-6047
ISBN 978-0-387-87707-5 e-ISBN 978-0-387-87708-2
DOI 10.1007/978-0-387-87708-2
Springer New York Dordrecht Heidelberg London
Library of Congress Control Number: 2010929771
Mathematics Subject Classification (2010): 37N25; 92C20; 92C05
c
Springer Science+Business Media, LLC 2010
All rights reserved. This work may not be translated or copied in whole or in part without the written
permission of the publisher (Springer Science+Business Media, LLC, 233 Spring Street, New York,
NY 10013, USA), except for brief excerpts in connection with reviews or scholarly analysis. Use in
connection with any form of information storage and retrieval, electronic adaptation, computer software,
or by similar or dissimilar methodology now known or hereafter developed is forbidden.
The use in this publication of trade names, trademarks, service marks, and similar terms, even if they are
not identified as such, is not to be taken as an expression of opinion as to whether or not they are subject
to proprietary rights.
Printed on acid-free paper
Springer is part of Springer Science+Business Media (www.springer.com)
10.
Preface
One can saythat the field of computational neuroscience started with the 1952 paper
of Hodgkin and Huxley in which they describe, through nonlinear partial differential
equations, the genesis of the action potential in the giant axon of the squid. These
equations and the methods that arose from this combination of modeling and ex-
periments have since formed the basis for nearly every subsequent model for active
cells. The Hodgkin–Huxleymodel and a host of simplified equations that are derived
from it have inspired the development of new and beautiful mathematics. Dynamical
systems and computational methods are now being used to study activity patterns
in a variety of neuronal systems. It is becoming increasingly recognized, by both
experimentalists and theoreticians, that issues raised in neuroscience and the math-
ematical analysis of neuronal models provide unique interdisciplinary collaborative
research and educational opportunities.
This book is motivated by a perceived need for an overview of how dynamical
systems and computational analysis have been used in understanding the types of
models that come out of neuroscience. Our hope is that this will help to stimulate
an increasing number of collaborations between mathematicians and other theo-
reticians, looking for interesting and relevant problems in applied mathematics and
dynamical systems, and neuroscientists, looking for new ways to think about the
biological mechanisms underlying experimental data.
The book arose out of several courses that the authors have taught. One of these
is a graduate course in computational neuroscience that has students from the dis-
ciplines of psychology, mathematics, computer science, physics, and neuroscience.
Of course, teaching a course to students with such diverse backgrounds presents
many challenges. However, the course provides many opportunities to encourage
students, who may not normally interact with each other, to collaborate on exercises
and projects. Throughout the book are many exercises that involve both computa-
tion and analysis. All of the exercises are motivated by issues that arise from the
biology.
We have attempted to provide a comprehensive introduction to the vocabulary
of neuroscience for mathematicians who are just becoming interested in the field,
but who have struggled with the biological details. Anyone who wants to work in
computational neuroscience should learn these details as this is the only way one
can be sure that the analysis and modeling is actually saying something useful to
v
11.
vi Preface
biologists. Wehighly recommend the reader study this material in more detail by
consulting one of the many excellent books devoted primarily to neuroscience. Such
books include those by Kandel et al. [144] and Johnston and Wu [139].
We have also tried to provide background material on dynamical systems theory,
including phase plane methods, oscillations, singular perturbations, and bifurcation
analysis. An excellent way to learn this material is by using it, together with com-
puter simulations, to analyze interesting, concrete examples. The only prerequisites
are a basic knowledge of calculus, knowledge of a little linear algebra (matrices,
eigenvalues), and understanding of some basic theory of ordinary differential equa-
tions. Much of the mathematics is at the level of Strogatz [255].
The book is organized from the bottom up. The first part of the book is con-
cerned with properties of a single neuron. We start with the biophysics of the cell
membrane, add active ion channels, introduce cable theory, and then derive the
Hodgkin–Huxley model. Chapter 2 is concerned with the basic properties of den-
drites. We then introduce dynamical systems theory, using a simple neuron model to
illustrate the basic concepts. We return to the biology in Chap. 4, where we discuss
the variety of ion channels which have been found in neurons. Chapters 5 and 6
are devoted to bursting oscillations and propagating action potentials, respectively.
Here, we use many of the dynamical systems techniques to describe mechanisms
underlying these behaviors. The second part of the book is concerned with neu-
ronal networks. In Chap. 7, we describe synaptic channels, which are the primary
way that neurons communicate with each other. Chapters 8 and 9 discuss two dif-
ferent approaches for studying networks. First, we assume weak coupling and use
phase-response methods. We then demonstrate how one can analyze firing patterns
in neuronal networks using fast/slow analysis. In Chap. 10, we discuss the role of
noise in neuron models. Here, we briefly introduce the reader to the mathematical
theory of stochastic differential equations. Finally, in Chaps. 11 and 12 we discuss
firing rate models and spatially distributed networks.
There is far more material in this book than could be covered in a one-semester
course. Furthermore, some of the material is quite advanced. A course in computa-
tional neuroscience slanted toward mechanisms and dynamics could easily be made
out of the first five chapters along with Chap. 7. These chapters would cover most
of the basics of single-cell modeling as well as introduce students to dynamical sys-
tems. The remainder of such a course could include selections from Chaps. 8–12.
For example, Chap. 11 contains firing rate models, with many applications provided
in Sect. 11.3. Parts of Chap. 12 could comprise the remainder of the course.
For more mathematically inclined students, the elementary dynamics chapter
(Chap.3) could be skipped and the more technical chapters could be emphasized.
There is lovely nonlinear dynamics in Chaps. 5, 6, 8, and 9, which along with the
earlier chapters could form the core of a mathematical neuroscience course.
There are several recent books that cover some of the same material as in
the present volume. Theoretical Neuroscience by Dayan and Abbott [53] has a
broader range of topics than our book; however, it does not go very deeply into
the mathematical analysis of neurons and networks, nor does it emphasize the
dynamical systems approach. A much more similar book is Dynamical Systems
12.
Preface vii
in Neuroscienceby Izhikevich [136]. This book emphasizes the same approach
as we take here; however, the main emphasis of Dynamical Systems in Neu-
roscience is on single-neuron behavior. We cover a good deal of single-neuron
biophysics, but include a much larger proportion of theory on systems neuro-
science and applications to networks. There are many specific models and equa-
tions in this text. The forms of these models and their parameters are available at
http://www.math.pitt.edu/bard/bardware/neurobook/allodes.html.
Pittsburgh, PA G. Bard Ermentrout
Columbus, OH David H. Terman
14.
Contents
1 The Hodgkin–HuxleyEquations ............................................ 1
1.1 The Resting Potential .................................................. 1
1.2 The Nernst Equation ................................................... 3
1.3 The Goldman–Hodgkin–Katz Equation............................... 5
1.4 Equivalent Circuits: The Electrical Analogue ........................ 8
1.5 The Membrane Time Constant ........................................ 11
1.6 The Cable Equation .................................................... 13
1.7 The Squid Action Potential ............................................ 16
1.8 Voltage-Gated Channels ............................................... 18
1.9 Hodgkin–Huxley Model ............................................... 20
1.10 The Action Potential Revisited ........................................ 25
1.11 Bibliography............................................................ 27
1.12 Exercises ................................................................ 28
2 Dendrites ...................................................................... 29
2.1 Multiple Compartments ................................................ 29
2.2 The Cable Equation .................................................... 33
2.3 The Infinite Cable ...................................................... 34
2.4 Finite and Semi-infinite Cables ........................................ 36
2.5 Branching and Equivalent Cylinders .................................. 38
2.6 An Isolated Junction ................................................... 40
2.7 Dendrites with Active Processes....................................... 42
2.8 Concluding Remarks ................................................... 45
2.9 Bibliography ............................................................ 45
2.10 Exercises ................................................................ 45
3 Dynamics ...................................................................... 49
3.1 Introduction to Dynamical Systems ................................... 49
3.2 The Morris–Lecar Model .............................................. 49
3.3 The Phase Plane ........................................................ 51
3.3.1 Stability of Fixed Points...................................... 52
3.3.2 Excitable Systems ............................................ 53
3.3.3 Oscillations ................................................... 55
ix
15.
x Contents
3.4 BifurcationAnalysis ................................................... 56
3.4.1 The Hopf Bifurcation......................................... 56
3.4.2 Saddle–Node on a Limit Cycle .............................. 58
3.4.3 Saddle–Homoclinic Bifurcation ............................. 60
3.4.4 Class I and Class II ........................................... 62
3.5 Bifurcation Analysis of the Hodgkin–Huxley Equations............. 63
3.6 Reduction of the Hodgkin–Huxley Model to a Two-Variable Model 66
3.7 FitzHugh–Nagumo Equations ......................................... 69
3.8 Bibliography ............................................................ 70
3.9 Exercises ................................................................ 70
4 The Variety of Channels ..................................................... 77
4.1 Overview................................................................ 77
4.2 Sodium Channels....................................................... 78
4.3 Calcium Channels ...................................................... 80
4.4 Voltage-Gated Potassium Channels ................................... 82
4.4.1 A-Current ..................................................... 83
4.4.2 M-Current ..................................................... 85
4.4.3 The Inward Rectifier.......................................... 86
4.5 Sag ...................................................................... 87
4.6 Currents and Ionic Concentrations .................................... 88
4.7 Calcium-Dependent Channels ......................................... 90
4.7.1 Calcium Dependent Potassium:
The Afterhyperpolarization .................................. 90
4.7.2 Calcium-Activated Nonspecific Cation Current............. 93
4.8 Bibliography ............................................................ 95
4.9 Exercises ................................................................ 95
4.10 Projects .................................................................100
5 Bursting Oscillations .........................................................103
5.1 Introduction to Bursting................................................103
5.2 Square-Wave Bursters..................................................105
5.3 Elliptic Bursting ........................................................111
5.4 Parabolic Bursting......................................................114
5.5 Classification of Bursters ..............................................117
5.6 Chaotic Dynamics ......................................................118
5.6.1 Chaos in Square-Wave Bursting Models ....................118
5.6.2 Symbolic Dynamics ..........................................121
5.6.3 Bistability and the Blue-Sky Catastrophe ...................123
5.7 Bibliography ............................................................125
5.8 Exercises ................................................................126
16.
Contents xi
6 PropagatingAction Potentials...............................................129
6.1 Traveling Waves and Homoclinic Orbits ..............................130
6.2 Scalar Bistable Equations ..............................................132
6.2.1 Numerical Shooting ..........................................135
6.3 Singular Construction of Waves .......................................136
6.3.1 Wave Trains ...................................................139
6.4 Dispersion Relations ...................................................139
6.4.1 Dispersion Kinematics .......................................141
6.5 Morris–Lecar Revisited and Shilnikov Dynamics ....................141
6.5.1 Class II Dynamics ............................................142
6.5.2 Class I Dynamics .............................................143
6.6 Stability of the Wave ...................................................145
6.6.1 Linearization ..................................................146
6.6.2 The Evans Function ..........................................147
6.7 Myelinated Axons and Discrete Diffusion ............................149
6.8 Bibliography ............................................................151
6.9 Exercises ................................................................152
7 Synaptic Channels ............................................................157
7.1 Synaptic Dynamics.....................................................158
7.1.1 Glutamate .....................................................161
7.1.2 ”-Aminobutyric Acid.........................................162
7.1.3 Gap or Electrical Junctions...................................164
7.2 Short-Term Plasticity...................................................164
7.2.1 Other Models .................................................167
7.3 Long-Term Plasticity ...................................................168
7.4 Bibliography ............................................................169
7.5 Exercises ................................................................169
8 Neural Oscillators: Weak Coupling ........................................171
8.1 Neural Oscillators, Phase, and Isochrons .............................172
8.1.1 Phase Resetting and Adjoints ................................174
8.1.2 The Adjoint ...................................................177
8.1.3 Examples of Adjoints.........................................178
8.1.4 Bifurcations and Adjoints ....................................181
8.1.5 Spike-Time Response Curves ................................186
8.2 Who Cares About Adjoints? ...........................................187
8.2.1 Relationship of the Adjoint and the Response
to Inputs .......................................................187
8.2.2 Forced Oscillators ............................................189
8.2.3 Coupled Oscillators...........................................193
8.2.4 Other Map Models............................................199
8.3 Weak Coupling .........................................................202
8.3.1 Geometric Idea................................................203
8.3.2 Applications of Weak Coupling..............................205
17.
xii Contents
8.3.3 SynapticCoupling near Bifurcations ........................206
8.3.4 Small Central Pattern Generators ............................208
8.3.5 Linear Arrays of Cells ........................................213
8.3.6 Two-Dimensional Arrays.....................................217
8.3.7 All-to-All Coupling...........................................219
8.4 Pulse-Coupled Networks: Solitary Waves ............................223
8.4.1 Integrate-and-Fire Model.....................................226
8.4.2 Stability .......................................................229
8.5 Bibliography ............................................................229
8.6 Exercises ................................................................229
8.7 Projects .................................................................238
9 Neuronal Networks: Fast/Slow Analysis ...................................241
9.1 Introduction.............................................................241
9.2 Mathematical Models for Neuronal Networks ........................242
9.2.1 Individual Cells ...............................................242
9.2.2 Synaptic Connections ........................................243
9.2.3 Network Architecture ........................................245
9.3 Examples of Firing Patterns ...........................................246
9.4 Singular Construction of the Action Potential ........................249
9.5 Synchrony with Excitatory Synapses..................................254
9.6 Postinhibitory Rebound ................................................258
9.6.1 Two Mutually Coupled Cells.................................258
9.6.2 Clustering .....................................................260
9.6.3 Dynamic Clustering ..........................................260
9.7 Antiphase Oscillations with Excitatory Synapses ....................262
9.7.1 Existence of Antiphase Oscillations .........................263
9.7.2 Stability of Antiphase Oscillations ..........................266
9.8 Almost-Synchronous Solutions........................................269
9.8.1 Almost Synchrony with Inhibitory Synapses ...............269
9.8.2 Almost Synchrony with Excitatory Synapses ...............271
9.8.3 Synchrony with Inhibitory Synapses ........................274
9.9 Slow Inhibitory Synapses ..............................................275
9.9.1 Fast/Slow Decomposition ....................................275
9.9.2 Antiphase Solution ...........................................276
9.9.3 Suppressed Solutions .........................................278
9.10 Propagating Waves .....................................................278
9.11 Bibliography............................................................282
9.12 Exercises ................................................................282
10 Noise............................................................................285
10.1 Stochastic Differential Equations......................................287
10.1.1 The Wiener Process ..........................................288
10.1.2 Stochastic Integrals ...........................................289
10.1.3 Change of Variables: Itô’s Formula ..........................289
18.
Contents xiii
10.1.4 Fokker–PlanckEquation: General Considerations ..........290
10.1.5 Scalar with Constant Noise ..................................293
10.1.6 First Passage Times...........................................295
10.2 Firing Rates of Scalar Neuron Models ................................299
10.2.1 The Fokker–Planck Equation ................................299
10.2.2 First Passage Times...........................................303
10.2.3 Interspike Intervals ...........................................306
10.2.4 Colored Noise.................................................307
10.2.5 Nonconstant Inputs and Filtering Properties ................309
10.3 Weak Noise and Moment Expansions.................................310
10.4 Poisson Processes ......................................................314
10.4.1 Basic Statistics ................................................314
10.4.2 Channel Simulations .........................................317
10.4.3 Stochastic Spike Models: Beyond Poisson ..................319
10.5 Bibliography ............................................................321
10.6 Exercises ................................................................321
10.7 Projects .................................................................326
11 Firing Rate Models ...........................................................331
11.1 A Number of Derivations ..............................................332
11.1.1 Heuristic Derivation ..........................................332
11.1.2 Derivation from Averaging...................................336
11.1.3 Populations of Neurons.......................................338
11.2 Population Density Methods...........................................341
11.3 The Wilson–Cowan Equations.........................................344
11.3.1 Scalar Recurrent Model ......................................346
11.3.2 Two-Population Networks ...................................346
11.3.3 Excitatory–Inhibitory Pairs ..................................350
11.3.4 Generalizations of Firing Rate Models ......................356
11.3.5 Beyond Mean Field...........................................359
11.4 Some Methods for Delay Equations...................................361
11.5 Exercises ................................................................363
11.6 Projects .................................................................365
12 Spatially Distributed Networks .............................................369
12.1 Introduction.............................................................369
12.2 Unstructured Networks ................................................370
12.2.1 McCulloch–Pitts ..............................................370
12.2.2 Hopfield’s Model .............................................371
12.2.3 Designing Memories .........................................373
12.3 Waves ...................................................................375
12.3.1 Wavefronts ....................................................376
12.3.2 Pulses..........................................................379
19.
xiv Contents
12.4 Bumps..................................................................383
12.4.1 The Wilson–Cowan Equations ...............................384
12.4.2 Stability .......................................................387
12.4.3 More General Stability .......................................388
12.4.4 More General Firing Rates ...................................389
12.4.5 Applications of Bumps .......................................390
12.5 Spatial Patterns: Hallucinations .......................................394
12.6 Exercises ................................................................399
References...........................................................................407
Index .................................................................................419
20.
Acknowledgments
I am deeplygrateful for the opportunity to have learned from and worked with
so many brilliant teachers, collaborators, and students. Thanks to them all. I espe-
cially thank Donald Aronson, for his encouragement to study mathematical biology,
Charles Conley, whose mathematical insights pervade this book, and John Rinzel,
for helping to pave the way into mathematical neuroscience. I would also like
to thank those who proofread and commented on earlier versions of the book.
These include Euiwoo Lee, Sungwoo Ahn, Choongseok Park, Michael Rempe,
Josic Kresmir, Robert McDougal, and Hossein Abbassian. Finally, thank you to the
National Science Foundation for its continued support. I dedicate the book to my
family Effat, Sarah, and Rochelle for their love, patience and support.
– David H. Terman, The Ohio State University, 2009
This book would not have been possible without the input of many students and
colleagues. Jack Cowan, Hugh Wilson, and John Rinzel had a keen appreciation for
how mathematics could provide insight into the working of the nervous system and,
as my teachers, they conveyed to me their knowledge and enthusiasm. I am grateful
for their efforts. I also want to thank the many students who took various versions
of courses I have taught on mathematical neuroscience and who provided valuable
feedback. In particular, I want to thank Jon Rubin, Brent Doiron, Sergio Verduzco,
and Kreso Jozic for comments on the many partial drafts of the book. Any mistakes
belong to me. I dedicate the book to my family, Ellen, Kyle, and Jordan, for their
patience with me over the years (and for putting up with the endless series of bad
puns and pointless stories). Finally, because I promised my nephew, B.S. King, I will
also dedicate the book to all the other “Bards” out there.
– G. Bard Ermentrout, The University of Pittsburgh, 2009
xv
22.
Chapter 1
The Hodgkin–HuxleyEquations
1.1 The Resting Potential
All living cells have an electrical voltage, or potential difference, between their
inside and outside. Since the cell’s membrane is what separates the inside from
the outside, this potential difference is referred to as the membrane potential. In
mathematical terms, the membrane potential VM is defined as
VM D Vin Vout;
where Vin is the potential on the inside of the cell and Vout is the potential on the
outside. This will change during an action potential, for example.
The resting potential refers to the potential across the membrane when the cell is
at rest. A typical neuron has a resting potential of about 70 mV. An inward current
corresponds to a positively charged ion, such as NaC
, entering the cell. This raises
the membrane potential; that is, it brings the membrane potential closer to zero. In
this case, the cell is said to be depolarized. An outward current corresponds to a
positively charged ion, such as KC
, leaving the cell or a negatively charged ion,
such as Cl
, entering the cell. In this case, the cell becomes hyperpolarized.
The potential difference arises from differences in the concentrations of various
ions within and outside the cell. The maintenance of the potential difference also
involves the transport of ions across the cell membrane and the selective permeabil-
ity of the membrane to these ions. The principal ions found on either side of the
cell membrane are NaC
, KC
, and Cl
. The concentration of KC
ions inside a cell
is about 10 times that in the extracellular fluid, whereas the concentrations of NaC
and Cl
are much higher outside the cell than inside.
The lipid bilayer of the cell membrane is a poor conductor of ionic current be-
cause it is not permeable to ions. However, the membrane does contain channel
proteins that allow for the ions to move through it. There are two types of ion chan-
nels in the membrane: gated and nongated. Nongated channels are always open,
whereas gated channels can open and close and the probability of opening often
depends on the membrane potential; these are referred to as voltage-gated chan-
nels. Gated channels are typically selective for a single ion. The permeability of the
G.B. Ermentrout and D.H. Terman, Mathematical Foundations of Neuroscience,
Interdisciplinary Applied Mathematics 35, DOI 10.1007/978-0-387-87708-2 1,
c
Springer Science+Business Media, LLC 2010
1
23.
2 1 TheHodgkin–Huxley Equations
membrane to a particular ion depends on the number of open channels selective for
that ion. Most gated channels are closed at rest; hence, the nongated ion channels
are primarily responsible for establishing the resting potential. An action potential
is generated when gated channels open allowing for the flux of ions across the cell
membrane.
Because of concentration differences, when the appropriate channels are open,
NaC
and Cl
ions tend to diffuse into the cell, whereas KC
ions tend to diffuse
outward. Note that ions do not simply diffuse in or out of an open channel until the
concentration of that ion on either side of the cell is zero. This is because of the
electric field created by separation of positive and negative charges across the cell
membrane.
Suppose, for example, the cell is permeable only to KC
. The concentration gra-
dient of KC
moves KC
ions out of the cell. However, the continued efflux of KC
builds up an excess of positive charge on the outside of the cell and leaves behind
an excess of negative charge on the inside. The negative charge consists mostly of
impermeable organic anions A
. This buildup of charge acts to impede the further
efflux of KC
, so eventually an equilibrium is reached. At this equilibrium, the elec-
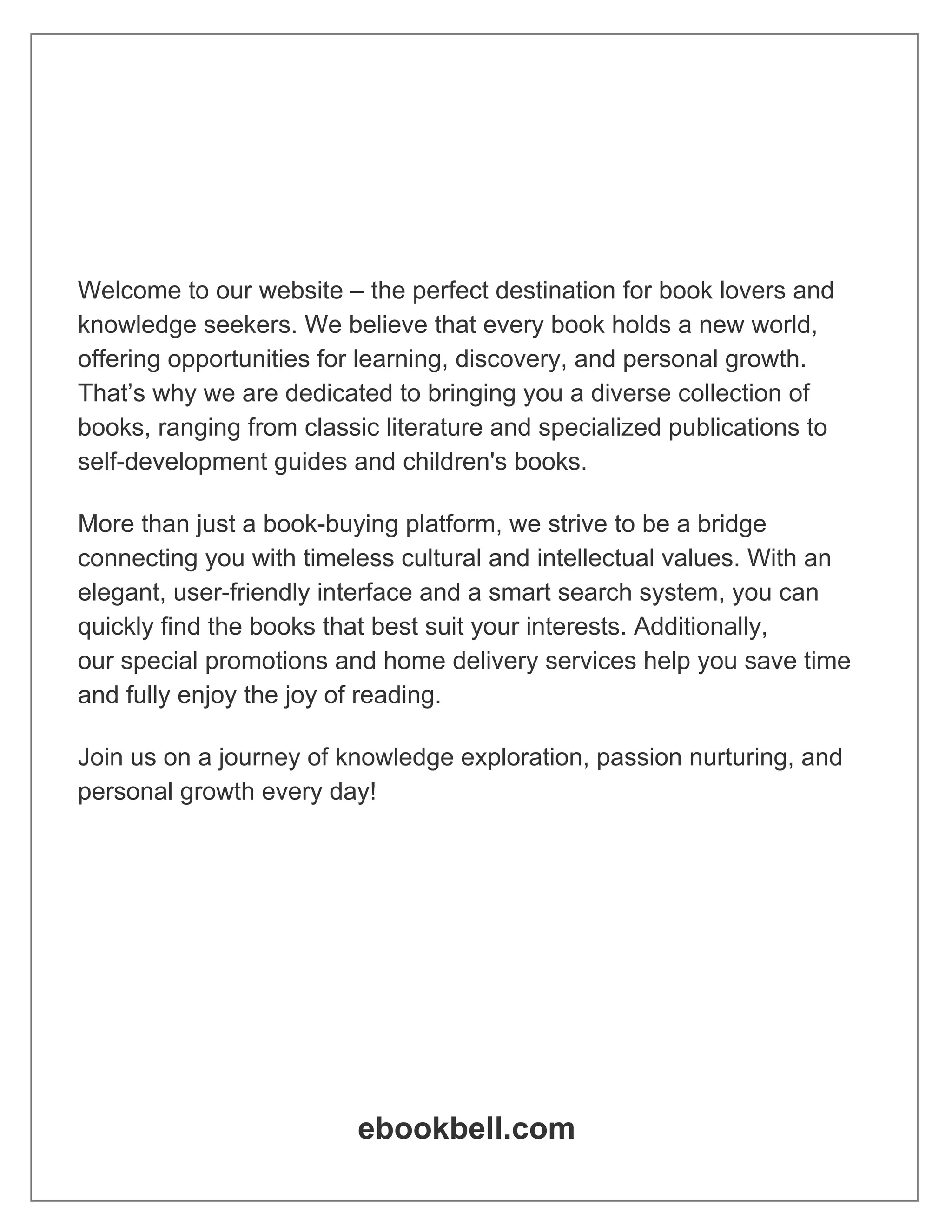
trical and chemical driving forces are equal and opposite (Fig. 1.1). The membrane
potential at which KC
ions are in equilibrium across the membrane is called the KC
Nernst, equilibrium, or reversal potential .
In the next section, we shall derive the following expression for the KC
Nernst
potential:
EK D
RT
zF
ln
ŒKC
in
ŒKCout
: (1.1)
A
−
A
−
A
−
A
−
+ + + + + + + + + + + +
- - - - - - - - - - - -
A
−
A
−
A
−
A
−
K+
K+
K+
K+
K+
K+
K+
K+
K+
K+
Na+
Na+
Na+ Na+
Na+
Na+
Na+
Na+
Cl
− Cl
−
Cl
−
Cl
−
a b
Extracellular side
Cytoplasmic side
Fig. 1.1 The KC
flux is determined by both the KC
concentration gradient and the electrical
potential across the membrane. (a) For a cell that is permeable only to KC
, the concentration
gradient of KC
moves KC
ions out of the cell. (b) The continued efflux of KC
builds up an excess
of positive charge on the outside and an excess of negative charge on the inside. At equilibrium,
the electrical and chemical driving forces are equal and opposite
24.
1.2 The NernstEquation 3
Here, EK is the KC
Nernst potential, R is the gas constant, T is the absolute
temperature in kelvin, z is the valence of KC
, F is Faraday’s constant, and ŒKC
out
and ŒKC
in are the concentrations of KC
ions outside and inside the cell. A similar
formula holds for the NaC
and Cl
Nernst potentials.
Neurons at rest are permeable to NaC
and Cl
in addition to KC
. Because of
their concentration differences, NaC
and Cl
ions move into the cell and KC
ions
move outward. The influx of NaC
ions tends to depolarize the cell, whereas the ef-
flux of KC
and the influx of Cl
have the opposite effect. The resting potential of
the cell is the potential at which there is a balance between these fluxes. It depends
on the concentrations of the ions both inside and outside the cell, as well as the per-
meability of the cell membrane to each of the ions. We note that at rest, many more
KC
and Cl
channels than NaC
channels are open; hence, the cell’s resting poten-
tial is determined primarily by the KC
and Cl
Nernst potentials. In the following
sections, we shall derive the Goldman–Hodgkin–Katz (GHK) equation, which gives
an explicit expression for how the resting potential depends on the concentrations,
both inside and outside, of ions and the permeabilities of the membrane to the ions.
For a cell to maintain a constant resting potential, the efflux of KC
ions must
balance the influx of NaC
ions (here we are ignoring Cl
ions). That is, the
charge separation across the membrane must be constant. If these steady ion leaks
continued unopposed, then KC
ions within the cell would become depleted, whereas
the concentration of NaC
ions inside the cell would increase. This would eventually
result in a loss of the ionic gradients, necessary for maintaining the resting poten-
tial. The dissipation of ionic gradients is prevented by active pumps that extrude
NaC
ions from the cell while taking in KC
. The NaC
–KC
pump is an integral mem-
brane protein that exchanges three NaC
ions for two KC
ions. This is probably the
most important ion transporter in biological membranes; however, there are many
other proteins in the membrane that are capable of pumping ions from one side of
the membrane to the other.
1.2 The Nernst Equation
Here we derive the Nernst equation and, in Sect. 1.3 we derive the GHK equation.
Recall that if the membrane is permeable to only one ion, then that ion’s Nernst
potential is the resting potential at which the electrical and chemical driving forces
balance. The GHK equation is, in some sense, a generalization of the Nernst equa-
tion in which we assume the membrane is permeable to more than just one ion. The
GHK equation determines the resting potential at which the electrical and chemical
forces, generated by each of these ions, balance each other. The first step in deriving
these equations is to derive the Nernst–Planck equation.
In what follows, let ŒC.x/ be the concentration of some ion and V.x/ the poten-
tial at the point x across the membrane. Then, Fick’s law of diffusion says that the
diffusive flux, Jdiff, is given by
Jdiff D D
@ŒC
@x
:
25.
4 1 TheHodgkin–Huxley Equations
The diffusion constant, D, has units of square centimeters per second and the
concentration is in molecules per cubic centimeter, so the diffusive flux has units of
molecules per square centimeter second. (Think of the flux as movement across the
two-dimensional cell surface.) The direction of movement is from high concentra-
tions to low concentrations. The diffusion constant (empirically measured) depends
on the size of the molecule and the medium in which it is diffusing. A typical value
for ions such as KC
, Cl
, and NaC
is 2:5 106
cm2
/s. Calcium ion has a diffusion
constant about an order of magnitude less.
The other physical force that is responsible for the passive movement of ions is
the electrical drift described by the microscopic version of Ohm’s law:
Jdrift D zŒC
@V
@x
:
The electric field, E @V =@x, is the gradient of the potential V (measured in
volts) and thus has units of volts per centimeter. z is the valence of the ion (˙1; ˙2;
etc.). The parameter is the mobility and has dimensions of square centimeters per
volt second and ŒC is the concentration. The higher the concentration, the greater
the drift. Note that the drift has the same dimensions as the diffusive flux.
The total flux across the membrane is given by the sum of the diffusive flux and
the electrical drift:
Jtotal D D
@ŒC
@x
zŒC
@V
@x
:
Einstein’s relation connects the mobility with the diffusion coefficient:
D D
kT
q
;
where k is Boltzmann’s constant (J/K), T is the absolute temperature, and q is the
charge (measured in coulombs). Thus, we can write the total flux as
Jtotal D
kT
q
@ŒC
@x
zŒC
@V
@x
:
It is convenient to convert this equation, which is in terms of the number of indi-
vidual molecules, into its molar equivalent, by dividing by Avogadro’s number. It
is also convenient to introduce RT=F , where R is the ideal gas constant and F is
Faraday’s constant, instead of kT=q. (A list of these constants is given at the end
of the next section.) This will yield the flux per mole. Multiplying this flux by the
valence and Faraday’s constant yields a current flux
I D
uzRT
@ŒC
@x
C uz2
F ŒC
@V
@x
measured in amperes per square centimeter. The quantity u is the molar mobility,
=NA: This equation is the Nernst–Planck equation.
26.
1.3 The Goldman–Hodgkin–KatzEquation 5
The Nernst equation is obtained by setting the current equal to zero. That is, for
a given ionic species, at equilibrium, the diffusion and electric effects balance:
I D
uzRT
@ŒC
@x
C uz2
F ŒC
@V
@x
D 0:
As an exercise, it is left to the reader to prove this implies the Nernst equation:
Veq Vin Vout D
RT
zF
ln
ŒCin
ŒCout
: (1.2)
That is, the equilibrium (or Nernst) potential, which occurs when all the fluxes bal-
ance, depends on the logarithm of the ratio of the concentrations of the ions inside
and outside the cell.
To illustrate how to use the Nernst equation to compute an equilibrium potential,
note that in a typical mammalian cell, there is 140 mM KC
inside the cell and 5 mM
outside. At room temperature, 37ı
C, RT=F D 26:73 mV. Hence, the equilibrium
potential of potassium is
62 log
140
5
D 89:7mV:
1.3 The Goldman–Hodgkin–Katz Equation
The Nernst–Planck equation describes the movement of charged ions in aqueous
media. However, the cell membrane has thickness and there may be energy barriers
or blocking sites within the channel. In this case, the ions flowing through the open
channel may not obey the Nernst–Planck equation and we must model the com-
plex behavior within the membrane to get a true picture of the flux across the cell.
This type of biophysics is beyond the details that are needed for this book, but
the resulting equation does play a role in later parts. Thus, we will present a short-
ened derivation of a simplification of what happens within the membrane. Goldman,
Hodgkin, and Katz came up with this simplified model called the constant-field
equation. They assumed (1) the electric field across the lipid membrane is constant,
(2) the Nernst–Planck equation holds within the membrane, and (3) the ions all
move independently.
Let VM be the total potential across a membrane of width l and let V.x/ be the
potential at the point x across the membrane. Since the electric field is constant,
E D VM=l. This implies that dV=dx D VM=l: The mobility of ions within the
membrane will be different from that in the aqueous solution; denote this mobility
by u
: Finally, let ˇ be the ratio of the ion solubility within the membrane to the ion
27.
6 1 TheHodgkin–Huxley Equations
solubility in the aqueous solution. Thus, if ŒC is the aqueous concentration, then
ˇŒC is the membrane concentration. With these assumptions, the Nernst–Planck
equation for current across the membrane is
I D u
z2
FˇŒC
VM
l
u
zRTˇ
dŒC
dx
; 0 x l:
This is just a first-order linear ordinary differential equation for ŒC subject to the
two boundary conditions
ŒC.0/ D ŒCin; ŒC.l/ D ŒCout:
One cannot, in general, solve a first-order equation with two boundary conditions.
However, the current I is unknown, so choosing this correctly will allow us to find
a solution that satisfies both boundary conditions. We leave this elementary exercise
for the reader. The result is
I D
u
z2
F VMˇ
l
ŒCoute
ŒCin
e 1
!
;
where
D
zVMF
RT
:
This expression is often written in terms of the permeability,
P
ˇu
RT
lF
I
that is,
I D P zF
ŒCoute
ŒCin
e 1
!
: (1.3)
The permeability has dimensions of centimeters per second. Thus, the dimensions
are in terms of current per unit area. Equation (1.3) is called the constant-field
equation.
This is the current due to a single ionic species. The current vanishes at the equi-
librium or Nernst potential of the ionic species. A current–voltage (I–V ) plot is a
common plot. If the inside and outside concentrations are identical, then the I–V
plot is linear. For ŒCout ŒCin (respectively, ŒCout ŒCin ) the I–V plot is con-
cave down (respectively concave up). The reader is encouraged to plot the current as
a function of the voltage for different concentration ratios. If the concentrations are
quite different on the inside and outside, then the I–V curve is strongly rectifying.
This means the magnitude of the current depends strongly on whether or not the
potential is above or below the equilibrium.
28.
1.3 The Goldman–Hodgkin–KatzEquation 7
Given several ionic species, the total current is just a sum of the individual cur-
rents. This is a consequence of assumption 3, which says that the ions do not interact.
Suppose there are three permeable ions, KC
, NaC
, and Cl
with corresponding
currents, IK; INa, and ICl: At equilibrium, the total current, I D IK C INa C ICl,
vanishes; that is, I D 0: The potential at which this occurs is
VM D
RT
F
ln
PKŒKC
out C PNaŒNaC
out C PClŒCl
in
PKŒKCin C PNaŒNaCin C PClŒClout
; (1.4)
where the Pj ’s are the permeabilities of each of the three ionic species. This is a
generalization of the Nernst equilibrium discussed above and is called the Goldman–
Hodgkin–Katz (GHK) equation. With one species, the equation reduces to the
Nernst potential. For example, in the squid axon, the ratios of the permeabilities,
at rest, are PK W PNa W PCl D 1 W 0:03 W 0:1. The ion concentrations inside the cell
are, respectively, for KC
, NaC
, and Cl
, 400, 50, and 40 mM, whereas outside the
cell they are 10, 460, and 540 mM. Thus, at room temperature, the equilibrium or
resting potential is 74 mV.
Table 1.1 Typical ion concentrations in cells (from Johnston and Wu [139])
Equilibrium potential (mV),
Ion Inside (mM) Outside (mM) Ei D RT
zF
ln
ŒCout
ŒCin
Frog muscle T D 20ı
C
KC
124 2.25 58 log 2:25
124
D 101
NaC
10.4 109 58 log 109
10:4
D C59
Cl
1.5 77.5 58 log 77:5
1:5
D 99
Ca2C
104
2.1 29 log 2:1
104 D C125
Squid axon T D 20ı
C
KC
400 20 58 log 20
400
D 75
NaC
50 440 58 log 440
50
D C55
Cl
40–150 560 58 log 560
40150
D 66 to 33
Ca2C
104
10 29 log 10
104 D C145
Mammalian cell T D 37ı
C
KC
140 5 62 log 5
140
D 89:7
NaC
5–15 145 62 log 145
515
D C90 .C61/
Cl
4 110 62 log 110
4
D 89
Ca2C
104
2.5–5 31 log 2:55
104 D C 136 .C145/
29.
8 1 TheHodgkin–Huxley Equations
Table 1.2 Elementary
constants
NA 6:022 1023
mol (Avogadro’s number)
k 1:380658 1023
J/K (Boltzmann’s constant)
R 8:31451 J/(mol K) (ideal gas constant)
e 1:602177 1019
C (electron charge)
F 96; 485:3 C/mol (Faraday’s constant)
0 8:85 1012
F/m (permittivity constant)
K Kelvin (degrees centigrade C273:16/
L Liter
N Newton
J Joule (N m); 1 J = 0:238845 cal
V Volt (J/C)
C Coulomb
A Ampere (C/s)
Ohm (V/A)
S Siemens (A/V)
F Farad (s A/V or C/V)
1.4 Equivalent Circuits: The Electrical Analogue
We saw in Sect. 1.3 that the electrical properties of cells are determined by the ionic
species that move through the membrane. Currents flow according to the permeabili-
ties of ion channels and concentration gradients across the cell membrane. However,
all of our discussion so far has been in a steady-state environment. The GHK equa-
tion does not determine how the membrane potential changes in response to changes
in the permeabilities. For this reason, it cannot be used to understand how these
changes in permeabilities may generate an action potential. A very useful way to
describe the behavior of the membrane potential is in terms of electrical circuits;
this is commonly called the equivalent circuit model. The circuit consists of three
components: (1) conductors or resistors, representing the ion channels; (2) batteries,
representing the concentration gradients of the ions; and (3) capacitors, represent-
ing the ability of the membrane to store charge. The equivalent circuit model leads
to both an intuitive and a quantitative understanding of how the movement of ions
generates electrical signals in the nerve cell.
We first consider a membrane that is only permeable to potassium. The equivalent
circuit is shown in Fig. 1.2. The lipid bilayer that constitutes the cell membrane has
dielectric properties and as such behaves in much the same manner as a capacitor.
Recall that capacitors store charge and then release it in the form of currents. The
relationship between the charge stored and the potential is given by
q D CMVMI (1.5)
that is, the total charge q is proportional to the potential VM with a proportional-
ity constant CM called the membrane capacitance. Note that the total capacitance
depends on the total area of the dielectric; thus, larger neurons have a larger total
capacitance than smaller ones. The capacitance per square centimeter is called
the specific membrane capacitance and will be denoted as cM. Hence, the total
30.
1.4 Equivalent Circuits:The Electrical Analogue 9
CM VM
RK
EK
Lipid
bilayer
Channel
Fig. 1.2 The cell membrane showing the insulating lipid bilayer and a KC
channel, which allows
current to flow. The equivalent electrical circuit is shown on the right
membrane capacitance CM is the specific membrane capacitance cM times the total
surface area of the cell. In general, the specific membrane capacitance may depend
on the potential; however, for most cell membranes, the specific membrane capaci-
tance is very close to 1 F/cm2
.
Since current is the time derivative of charge, we can differentiate (1.5), divide
by the cell’s area, and obtain an expression for the specific capacitance current:
icap D cM
dVM
dt
: (1.6)
This gives the capacitance current per unit area. We will denote the total capacitance
current as Icap.
In the equivalent circuit, KC
channels are represented as a conductor in series
with a battery. If O
gK is the conductance of a single KC
channel, then, using Ohm’s
law, the ionic current through this channel is
O
IK D O
gK.VM EK/: (1.7)
Here, EK is the potential generated by the battery; this is given by the KC
Nernst
potential. The driving force is VM EK. Now suppose there are NK KC
channels in a
unit area of membrane. These can all be combined into the single equivalent circuit
shown in Fig. 1.2. The conductance per unit area, or specific membrane conductance
(S/cm2
), is given by gK D NK O
gK and the specific membrane resistance ( cm2
) is
rK 1=gK. Since the Nernst potential depends only on the concentration gradient
of KC
, and not on the number of KC
channels, it follows that the KC
current, per
unit area, is given by
IK D gK.VM EK/ D
VM EK
rK
: (1.8)
Kirchhoff’s current law states that the total current into the cell must sum to
zero. Together with the equivalent circuit representation, this leads to a differential
equation for the membrane potential:
0 D icap C IK D cM
dVM
dt
C
VM EK
rK
(1.9)
31.
10 1 TheHodgkin–Huxley Equations
Fig. 1.3 Equivalent circuit
for a membrane with three
channels
EK ENa
gCl
ECl
CM
gK gNa
I(t)
or
cM
dVM
dt
D
VM EK
rK
D gK.VM EK/: (1.10)
Figure 1.3 shows an equivalent circuit with three parallel conductances and a
current source, I.t/. Here the capacitance current must be equal to the sum of the
ionic currents and the current source. As before, the capacitance current, per unit
area, is given by (1.6) and the ionic current, per unit area, is given by
iion D gCl.VM ECl/ gK.VM EK/ gNa.VM ENa/: (1.11)
The current source is not typically expressed as current per unit area, so we must
divide I.t/ by the total surface area of the neuron, A. It then follows that
cM
dVM
dt
D gCl.VM ECl/ gK.VM EK/ gNa.VM ENa/ C I.t/=A: (1.12)
Note that we can rewrite this equation as
cM
dVM
dt
D
.VM ER/
rM
C I.t/=A; (1.13)
where
ER D .gClECl C gKEK C gNaENa/rM
is the cell’s resting potential and
rM D
1
gCl C gK C gNa
is the specific membrane resistance.
For a passive membrane in which the conductances and currents are all constant,
VM will reach a steady state:
Vss D
gClECl C gKEK C gNaENa C I=A
gCl C gk C gNa
:
32.
1.5 The MembraneTime Constant 11
In the absence of the applied current, the steady-state potential is a weighted sum of
the equilibrium potentials of the three currents. This is similar to the GHK equation
(1.4), in which the contribution to the resting potential by each ion is weighted in
proportionto the permeability of the membrane to that particular ion. Note, however,
that in the equivalent circuit model, the equilibrium is a linear weighted sum of the
equilibrium potentials, whereas in the GHK equation, the sum is nonlinear.
We remark that membrane conductance and permeability are related concepts;
however, they are not the same. The permeability depends on the state of the mem-
brane, whereas conductance depends on both the state of the membrane and the
concentration of the ions. The permeability to KC
, for example, may be high if
there are a large number of open KC
channels. However, if the concentration of KC
ions is low on both sides of the membrane, then the KC
conductance will be low.
1.5 The Membrane Time Constant
In this section, we consider how a passive, isopotential cell responds to an applied
current. This will help explain how each component of the electrical circuit con-
tributes to changes in the membrane potential. The cell is said to be passive if its
electrical properties do not change during signaling. Such a cell cannot generate
an action potential; however, it is important to understand how a cell’s passive, or
constant, properties influence changes in the membrane potential before consider-
ing active signaling. Moreover, many dendrites do not have gated channels, so their
behavior is influenced primarily by their passive properties. The cell is said to be
isopotential if the membrane potential is uniform at all points of the cell; that is,
the membrane potential depends only on time. To simplify the analysis, we will
consider a spherical cell with radius .
Suppose this cell is injected with an applied current, I.t/, that is turned on at
t D 0 to some constant value, I0, and turned off at t D T . Here, we assume I0 0;
however, this is really not necessary. Note that for an isopotential cell, the injected
current distributes uniformly across the surface. It follows that for a spherical cell,
the current flowing across a unit area of the membrane is
IM.t/ D
I.t/
4 2
D
8
ˆ
:̂
I0
42 if 0 t T
0 otherwise:
(1.14)
As before, suppose cM is the specific membrane capacitance, rM is the specific
membrane resistance, and ER is the cell’s resting potential. To simplify things, we
take ER D 0 so that VM measures the deviation of the membrane potential from rest.
From (1.13), the membrane potential satisfies the ordinary differential equation
cM
dVM
dt
D
VM
rM
C IM.t/: (1.15)
33.
12 1 TheHodgkin–Huxley Equations
Fig. 1.4 The change
of membrane potential in
response to a step of current.
The membrane potential is
shown with a solid line. The
dashed lines show the time
courses of the purely
capacitive and resistive
elements. The bottom panel
shows the time course of the
total membrane current, the
ionic current, and the
capacitive current
VM
purely
capacitive
purely
resistive
ImRM
time
IM
Im
VM
Iion
Iion
Icap
Icap
IM
If the cell starts at rest, then the solution of this linear equation satisfies
VM.t/ D
rMI0
4 2
1 e
t
M
for 0 t T; (1.16)
where M cMrM is the membrane time constant and
VM.t/ D VM.T /e t
M for t T: (1.17)
The solution is shown in Fig. 1.4. Once the current is turned on, the membrane
potential asymptotically approaches the steady-state value rMI0=.4 2
/. The ap-
proach is exponential with the time constant M. The membrane time constant also
determines the rate at which the membrane potential decays back to rest after the
current is turned off. The steady-state membrane potential satisfies
I0
rM
4 2
I0RINP; (1.18)
where RINP is the input resistance of the cell. Note that if the input current changes
by I, then the steady-state membrane potential changes by RINP I; that is, the
input resistance is the slope of the I–V curve obtained by plotting the steady-state
voltage against the injected current.
The initial rise in membrane potential is determined primarily by the membrane
capacitance. Initially, the voltage across the resistor and that across the capacitor
are both zero. From Ohm’s law, it follows that initially no current flows through the
resistor and all the current is due to the capacitor. Because of the capacitive current,
the potential across the capacitor, and hence the membrane potential, will become
34.
1.6 The CableEquation 13
more positive. As VM increases, the membrane potential difference begins to drive
current across the membrane resistance, resulting in less current across the capacitor.
Eventually, the membrane potential reaches a value where all the membrane current
flows through the resistor. This value is given by VM D I0RINP.
Figure 1.4 also shows responses in which there are purely resistive or purely
capacitive elements. If there is no membrane capacitance, then VM satisfies
VM.t/ D rMIM.t/: (1.19)
That is, VM jumps to the steady-state potential, I0RINP, as soon as the injected cur-
rent is turned on and it jumps back to rest as soon as the current is turned off. If
there is only a capacitive element, then the membrane potential changes linearly as
long as there is an applied current.
1.6 The Cable Equation
We have, so far, considered the passive properties of an isopotential cell. This
analysis may be used to describe signaling within the cell body, which can be ap-
proximated by a sphere. However, it is clearly not appropriate for studying electrical
properties of the axon or dendrites. These are better approximated by cylinders that
are not isopotential. A subthreshold voltage signal that is initiated at one point along
the axon or dendrite will decrease in amplitude with distance from the point of initi-
ation. It is important to understand how the geometry of the cell affects the spread of
the signal. The signal may, for example, correspond to synaptic input from another
neuron. Understanding how geometry affects the spread of the signal will help deter-
mine whether the synaptic input will cause the cell to fire an action potential. Here,
we assume the membrane is passive, so the analysis is more applicable to dendrites
than to axons. However, as we shall describe later, the passive spread of current flow
helps determine the velocity of propagating action potentials in the axon.
We consider a cell that is shaped as a long cylinder, or cable, of radius a. We
assume the current flow is along a single spatial dimension, x, the distance along
the cable. In particular, the membrane potential depends only on the x variable,
not on the radial or angular components. The cable equation is a partial differential
equation that describes how the membrane potential VM.x; t/ depends on currents
entering, leaving, and flowing within the neuron. The equivalent circuit is shown in
Fig. 1.5. In what follows, we will assume Re D 0, so that the extracellular space is
isopotential. This assumption is justified if the cable is in a bath with large cross-
sectional area.
We first consider the axial current flowing along the neuron due to voltage gra-
dients. Note that the total resistance of the cytoplasm grows in proportion to the
length of the cable and is inversely proportional to the cross-sectional area of the
cable. The specific intracellular resistivity, which we denote as rL, is the constant of
proportionality. Hence, a cable of radius a and length x has a total resistance of
35.
14 1 TheHodgkin–Huxley Equations
x
RL
Re
CM
RM
Ilong
a
IM
Fig. 1.5 Equivalent circuit for a uniform passive cable. Ilong is the current along the inside of
the cable, IM is the current across the membrane, RL is the resistance of the cytoplasm, Re is the
resistance of the extracellular space, RM is the membrane resistance, and CM is the membrane
capacitance
RL D rL x=. a2
/. It follows from Ohm’s law that at any point x, the decrease in
VM with distance is equal to the current times the resistance. That is,
VM.x C x; t/ VM.x; t/ D Ilong.x; t/RL D Ilong.x; t/
x
a2
rL: (1.20)
There is a minus sign because of the convention that positive current is a flow of
positive charges from left to right. If voltage decreases with increasing x, then the
current is positive. In the limit x ! 0,
Ilong.x; t/ D
a2
rL
@VM
@x
.x; t/: (1.21)
Let iion be the current per unit area due to ions flowing into and out of the cell.
Then the total ionic current that flows across a membrane of radius a and length x
is given by Iion D .2 a x/iion.
Recall that the rate of change of the membrane potential is determined by
the capacitance. The total capacitance of a membrane is equal to the specific
membrane capacitance cM multiplied by the total surface area of the membrane.
Hence, for a cable of radius a and length x, the total capacitance is given by
CM D .2 a x/cM and the amount of current needed to change the membrane
potential at a rate @VM=@t is
Icap.x; t/ D .2 a x/cM
@VM
@t
: (1.22)
From Kirchhoff’s law, the change in intracellular axial current is equal to the
amount of current that flows across the membrane. Hence,
Icap.x; t/ C Iion.x; t/ D Ilong.x C x; t/ C Ilong.x; t/; (1.23)
36.
1.6 The CableEquation 15
from which it follows that
.2 a x/cM
@VM
@t
C .2 a x/iion D
a2
rL
@VM
@x
.x C x; t/
a2
rL
@VM
@x
.x; t/:
We divide both sides of this equation by 2 a x and let x ! 0 to obtain the cable
equation:
cM
@VM
@t
D
a
2rL
@2
VM
@x2
iion: (1.24)
For a passive cable, in which the resting potential is assumed to be zero,
iion D VM.x; t/=rM; (1.25)
where rM is the specific membrane resistance. Then (1.24) becomes
cM
@VM
@t
D
a
2rL
@2
VM
@x2
VM
rM
: (1.26)
We can rewrite this equation as
M
@VM
@t
D 2 @2
VM
@x2
VM; (1.27)
where
D
r
arM
2rL
and M D cMrM (1.28)
are the space or length constant and the membrane time constant, respectively. Note
that the space constant depends on the geometry of the cable, that is, the cable’s
diameter; however, the time constant does not.
Later, we shall give a detailed analysis of solutions to the cable equation and
properties of passive dendrites. For now, it is instructive to consider steady-state so-
lutions. Suppose, for example, we consider a semi-infinite cable (defined for x 0)
and we inject a step of current, I0, at x D 0. As t ! 1, the solution VM.x; t/
approaches a steady-state solution Vss.x/ that does not depend on time. Setting
@VM
@t D 0 in (1.27), we find that Vss satisfies
2 d2
Vss
dx2
Vss D 0: (1.29)
To solve this equation, we need boundary conditions. Recall from (1.21) that
I0 D
a2
rL
@VM
@x
:
37.
16 1 TheHodgkin–Huxley Equations
It follows that Vss must satisfy the boundary condition
dVss
dx
.0/ D
rL
a2
I0: (1.30)
The solution of (1.29) and (1.30) is
Vss.x/ D
rL
a2
I0ex=
: (1.31)
Note that the membrane potential decays exponentially. The distance at which
the potential has decayed to 1=e is the space constant . Since the space constant
is proportional to the square root of the cable’s radius, we conclude that thicker
axons or dendrites have larger space constants than narrower processes. That is,
thicker processes transmit signals for greater distances. As we discuss later, this
is important because it influences the ability of the neuron to spatially summate
incoming synaptic potentials. Moreover, the electrotonic, or passive, conductance
plays an important role in the propagation of the action potential. Thicker cells with
a larger space constant are more easily excited and are able to generate faster action
potentials.
The input resistance is defined to be the steady-state membrane potential, evalu-
ated at x D 0, divided by the injected current. That is,
Rinp D Vss.0/=I0 D
rL
a2
D
1
a3=2
p
rMrL=2: (1.32)
Note that the input resistance of the cable varies with the 3=2 power of the cable
radius. Therefore, the input conductance is directly proportional to the 3=2 power of
the cable radius. The input resistance is important because it is something that can be
measured experimentally. Since it is also possible to measure the space constant ,
one can compute rM and rL from experimental data.
1.7 The Squid Action Potential
We have so far viewed the membrane as a passive cable. However, linear cables
cannot transmit information over long distances unless the cable has an enormous
diameter. For example, the squid axon is more than 5 cm long, has a diameter
of about a 0.5 mm, a resting membrane resistance of rM D 700 cm2
, and a
transmembrane resistance of rL D 30 cm: Thus, the space constant for the squid
axon is D 5:4 mm: This is an order of magnitude smaller than the length. If the
potential at one end of the axon is held at 120 mV above rest, then the potential at
the other end is about 10 V above the rest, a 10,000-fold decrement. For neural
signals to reach any distance, there must be another way to carry them so that they
do not degrade.
38.
1.7 The SquidAction Potential 17
Fig. 1.6 Equivalent circuit
underlying the
Hodgkin–Huxley equations
EK ENa
gL
EL
CM
gK gNa
I(t)
Nature has solved this problem by inserting voltage-gated channels into the
membranes of many cell types. These channels are proteins which selectively let
different ionic species into the cell. Furthermore, the permeability of the channels
depends on the local environment near the channel. In particular, for voltage-gated
channels, whether the channel is open or closed depends on the local potential near
the channel. It is the opening and closing of voltage-gated channels that is responsi-
ble for the generation of the action potential that propagates along the axon.
Hodgkin and Huxley (1952) were the first to provide a comprehensive, quanti-
tative description of the regenerative currents generating the action potential. The
choice of the squid axon was fortuitous since the electrical properties rely primarily
on NaC
and KC
ions. Consider the equivalent circuit shown in Fig. 1.6 and assume
the cell is isopotential. Then the membrane potential satisfies
cM
dV
dt
D gNa.V ENa/ gK.V EK/ gL.V EL/:
Here, we write V instead of VM and IL gL.V EL/ is called the leak current.
It corresponds to passive flow of ions through nongated channels. The leak conduc-
tance, gL, is constant. Since most nongated channels are permeable to KC
ions, EL
is close to EK. The conductances gNa and gK may change with time since these cor-
respond to the opening and closing of NaC
and KC
channels, respectively. At rest,
gK is about 30-fold bigger than gNa, so the resting state is near EK at about 65 mV.
Suppose we could increase the conductance of gNa 100-fold, then the resting poten-
tial would be much closer to the Nernst potential of NaC
, which is about C55 mV.
Thus, the amplification of the potential, such as during an action potential, involves
changes in the relative conductances of the dominant ionic species. Hodgkin and
Huxley’s insight was that voltage-gated channels provide the substrate for this dy-
namic regulation of the conductances.
The basic mechanisms underlying action potentials are the following (Fig. 1.7).
At rest, most of the NaC
channels are closed, so the membrane potential is deter-
mined primarily by the KC
Nernst potential. If the cell is depolarized above some
threshold, then NaC
channels open and this further depolarizes the cell. This allows
39.
18 1 TheHodgkin–Huxley Equations
Na+
channels
close
Na+
channels
open
K+
channels
open
Refractory period
EK
ENa
Vrest
V
time
Fig. 1.7 The action potential. During the upstroke, NaC
channels open and the membrane po-
tential approaches the NaC
Nernst potential. During the downstroke, NaC
channels are closed,
KC
channels are open, and the membrane potential approaches the KC
Nernst potential
even more NaC
channels to open, allowing more NaC
ions to enter the cell and
forcing the cell toward the NaC
Nernst potential. This is the upstroke of the ac-
tion potential. The NaC
channel is transient, so even when they are depolarized,
the NaC
channels eventually shut down. In the meantime, the depolarization opens
KC
channels and KC
ions exit the cell. This hyperpolarizes the cell as the mem-
brane potential moves toward the KC
equilibrium potential. Until the voltage-gated
KC
channels close up again, the membrane is refractory. During this time, pumps
exchange excess NaC
ions inside the cell with excess KC
ions outside the cell.
Only a very small change in the concentration of NaC
ions is needed to generate
an action potential. From the exercises, we find that approximately 53 million NaC
ions must diffuse across the membrane to depolarize it from 60 to C50 mV. This
influx of NaC
ions represents only a 0.012% change in the internal NaC
concentra-
tion, which is typically around 12 mM. Hence, changes in local charge separation,
not in concentration, are required for an action potential.
1.8 Voltage-Gated Channels
In the Hodgkin–Huxley model, each channel is viewed as a transmembrane protein
that forms a pore through which ions can diffuse down their concentration gradi-
ents. The pores have gates that can be either open or closed; the probability that a
40.
1.8 Voltage-Gated Channels19
gate is open or closed depends on the membrane potential. The gate model can be
summarized by the diagram
C
˛.V /
•
ˇ.V /
O; (1.33)
where C and O correspond to the closed and open states, respectively, and ˛.V /
and ˇ.V / are the voltage-dependent rate constants at which a gate goes from the
closed to the open and from the open to the closed states, respectively. If we let m
be the fraction of open gates, then 1 m is the fraction of closed gates, and, from
the law of mass action,
dm
dt
D ˛.V /.1 m/ ˇ.V /m D .m1.V / m/= .V /; (1.34)
where
m1.V / D
˛.V /
˛.V / C ˇ.V /
and .V / D
1
˛.V / C ˇ.V /
: (1.35)
It is easy to solve this equation if V is constant. The solution starting at m.0/ is
m.t/ D m1.V / C .m.0/ m1.V //et=.V /
:
Note that the solution approaches the steady-state m1.V / at a rate determined by
the time constant .V /.
One must obtain expressions for the voltage-dependent rate constants ˛ and ˇ.
In the Hodgkin–Huxley model, these functions were derived by fitting the data.
Borg-Graham [17] and others have suggested a simple formulation based on thermo-
dynamics. The idea is that the probability of opening or closing a channel depends
exponentially on the potential. Thus,
˛.V / D A˛ exp.B˛V / and ˇ.V / D Aˇ exp.Bˇ V /: (1.36)
From this, we find that
m1.V / D
1
1 C exp..V Vh/=Vs/
;
where Vh and Vs are constants. We leave as an exercise the calculation of these
constants in terms of the constants A and B. The time constant, .V /, will generally
be a skewed bell-shaped function of V: If Bˇ D B˛, then .V / is a hyperbolic
secant.
41.
20 1 TheHodgkin–Huxley Equations
1.9 Hodgkin–Huxley Model
We are now ready to derive the Hodgkin–Huxley model for the propagation of an
action potential along the squid’s giant axon. We view the axon as a cylinder of fixed
radius, a, so the membrane potential depends on the spatial variable x and time t.
Here, we assume there are voltage-gated KC
and NaC
channels and a leak current.
Then balancing currents, as in (1.23), we have
IL D Icap C Iion (1.37)
or, using (1.6) and (1.24),
a
2rL
@2
VM
@x2
D cM
@VM
@t
C IK C INa C IL: (1.38)
If each ionic current is ohmic, then this can be written as
cM
@VM
@t
D
a
2rL
@2
VM
@x2
gK.VM EK/ gNa.VM ENa/ gL.VM EL/: (1.39)
To complete the model, we need to describe how one computes the membrane con-
ductances gK; gNa, and gL. Note that the voltage-gated conductances gK and gNa
change with time during an action potential.
Hodgkin and Huxley used two experimental methods to separate the ionic cur-
rents and compute how the KC
and NaC
conductances depend on voltage. The first
was a simple feedback circuit called the voltage clamp that allows the experimenter
to hold the membrane potential at a constant or holding level VC. The voltage clamp
does so by injecting a current into the axon that is equal and opposite to the cur-
rent flowing through the voltage-gated channels. Electrical details can be found in
the book by Johnston and Wu [139]. Note that the voltage clamp separates the total
membrane current into its ionic and capacitive components. Recall that the capaci-
tive current satisfies Icap D CMdVM=dt. If the membrane potential is fixed at some
constant, then the capacitive current must be zero. Moreover, the total current can be
made spatially uniform by inserting a highly conductive axial wire inside the fiber;
the axon is then said to be space-clamped. In this case, @2VM
@x2 D 0: It then follows
that any changes in current must be due to either the leak or the opening and closing
of voltage-gated membrane channels.
We first consider how the voltage clamp can be used to determine the leak
conductance, gL. Note that most of the voltage-gated channels are closed at rest.
Moreover, if we hyperpolarize the cell, then we may assume all of the voltage-gated
channels are closed. It follows that if the membrane potential is clamped at some
sufficiently strong hyperpolarized level, then the total current is given by the leak;
that is,
IM gL.VC EL/:
From this equation, we can easily solve for gL.
42.
1.9 Hodgkin–Huxley Model21
Fig. 1.8 Numerically
computed voltage-clamp
experiment. The membrane
potential is stepped from rest
to 0 mV. This results in an
inward current followed by an
outward current. The separate
KC
and NaC
currents are
also shown
0 1 2 3 4 5 6
time (msec)
IK
IM
INa
VC = 0 mV
Figure 1.8 shows the results of a (numerically computed) voltage-clamp experi-
ment when the membrane potential is clamped at 0 mV. Note that there is an inward
current followed by an outward current. This result suggests the depolarizing volt-
age step turns on two voltage-gated channels. The inward current is due to the influx
of NaC
ions, whereas the outward current is due to the outward flow of KC
ions. It
is not clear, however, how these two separate ions contribute to the total membrane
current. For this it is necessary to isolate the two voltage-gated currents.
Hodgkin and Huxley were able to isolate the KC
current by replacing NaC
ions
in the external bathing solution with a larger, impermeant cation. This eliminated
the inward NaC
current. Now there are dozens of compounds that selectively block
different currents, many derived from natural toxins. (For example, tetrodotoxin,
which blocks NaC
channels, comes from the Pacific puffer fish, a tasty, if slightly
dangerous, Japanese delicacy called fugu.) Once NaC
has been removed, the voltage
clamp can be used to determine how IK depends on the membrane potential. That is,
one holds the membrane potential at various levels and determines the time course of
the total membrane current IM. If NaC
is removed, then the KC
current is computed
by subtracting the leak current from IM.
It is also now possible to block KC
channels using the drug tetraethylammonium.
This was not available to Hodgkin and Huxley; however, if IK and IL are known,
then one computes INa simply by subtracting IK and IL from IM. Once these cur-
rents have been determined, we can calculate the IK and INa conductances using
Ohm’s law. That is,
gK.t/ D
IK.t/
.VM EK/
and gNa.t/ D
INa.t/
.VM ENa/
: (1.40)
Figure 1.9 shows the IK and INa conductances for different levels of the holding
potential. Note than gNa turns on more rapidly that gK. Moreover, the NaC
channels
43.
22 1 TheHodgkin–Huxley Equations
−60
−40
−20
0
20
40
V
C
(mV)
0
10
20
30
0 1 2 3 4 5 6 7
0
20
40
time (msec)
gK
gNa
Fig. 1.9 Numerically computed voltage-clamp experiment. The membrane potential is stepped to
different values and the resulting KC
and NaC
conductances are computed
begin to close before the depolarization is turned off, whereas the KC
channels
remain open as long as the membrane is depolarized. This suggests the NaC
channel
can exist in three states: resting, activated, and inactivated. When the cell is depo-
larized, the NaC
channels switch from the resting (closed) to the activated (open)
state. If the depolarization is maintained, then the channel switches to the inactivated
(closed) state.
A physical interpretation of the NaC
channel is shown in Fig. 1.10. There are
two gates in the NaC
channel: a fast one (the activation gate), represented by the
line, and a slow one (the inactivation gate), represented by the ball. Both gates must
be open for the channel to conduct NaC
ions. At rest, the activation gate is closed
and the inactivation gate is open. When the membrane is depolarized, the activation
gate opens, which allows NaC
into the cell. The inactivation gate (ball) closes at
the higher potential, so the flow of NaC
is transient. Hodgkin and Huxley used a
more complicated voltage-clamp protocol, first stepping to a fixed voltage and then
applying brief voltage steps to probe the fast activation and slow inactivation gates.
Details can be found in [144].
Using the voltage-clamp data, Hodgkin and Huxley derived expressions for the
KC
and NaC
conductances. They proposed that
gK D N
gKn4
and gNa D N
gNam3
h; (1.41)
where N
gK and N
gNa are maximum conductances and n, m, and h are gating variables
that take values between 0 and 1. Hence, n4
represents the probability that a KC
44.
1.9 Hodgkin–Huxley Model23
Fig. 1.10 The
Hodgkin–Huxley NaC
channel. (a–c) Voltage-clamp
dynamics. (d) Physical model
of the channel. If the voltage
step is small (d, top) then the
NaC
channel’s activation gate
(line) is closed but the
inactivation gate (ball) is
open. At intermediate steps
(d, middle), both gates are
partially open. For large steps
(d, bottom), the activation
gate is open and the
inactivation gate is closed
Vrest
INa
V(t)
a
b
c
d
Vrest
channel is open: the KC
channel has four independent components, all of which
are identical. The probability that the sodium activation gate is open is m3
and the
probability that the sodium inactivation gate is open is h. Each of the gating variables
satisfies a first-order differential equation of the form (1.34). That is, they satisfy
equations of the form
dn
dt
D ˛n.V /.1 n/ ˇn.V /n D .n1.V / n/= n.V /;
dm
dt
D ˛m.V /.1 m/ ˇm.V /m D .m1.V / m/= m.V /;
dh
dt
D ˛h.V /.1 h/ ˇh.V /h D .h1.V / h/= h.V /:
If X D n, m, or h, then
X1.V / D
˛X .V /
˛X .V / C ˇX .V /
and X .V / D
1
˛X .V / C ˇX .V /
: (1.42)
To match the data, Hodgkin and Huxley chose the following parameters and gat-
ing functions: N
gNa D 120 mS=cm3
, N
gK D 36 mS=cm3
, N
gL D 0:3 mS=cm3
,
ENa D 50 mV; EK D 77 mV; EL D 54:4 mV;
˛n.V / D 0:01.V C 55/=.1 exp..V C 55/=10//;
ˇn.V / D 0:125 exp..V C 65/=80/;
˛m.V / D 0:1.V C 40/=.1 exp..V C 40/=10//;
45.
24 1 TheHodgkin–Huxley Equations
ˇm.V / D 4 exp..V C 65/=18/;
˛h.V / D 0:07 exp..V C 65/=20/;
ˇh.V / D 1=.1 C exp..V C 35/=10//:
In Fig. 1.11, we plot the activation curves n1.V /; m1.V /, and h1.V / along
with n.V /; m.V /, and h.V /. Note that n1 and m1 are increasing functions that
approach 0 for hyperpolarizing currents and approach 1 for depolarizing currents.
Hence, n and m become activated when the membrane is depolarized. On the other
hand, h1.V / is a decreasing function, so the NaC
channels inactivate when the
membrane is depolarized. It is also important to note that m.V / is considerably
smaller than n or h. Hence, NaC
channels activate much faster than they inactivate
or KC
channels open. In Fig. 1.12, we show the response of m, h, and n to a step in
voltage.
τ
(msec)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
−100 −80 −60 −40 −20 0 20 40
0
1
2
3
4
5
6
7
8
9
−100 −80 −60 −40 −20 0 20 40
V (mV)
steady
state
m
n
h
m
h
n
V (mV)
Fig. 1.11 Hodgkin–Huxley functions. Left the steady-state opening of the gates and right the time
constants
0 1 2 3 4 5 6
0
0.2
0.4
0.6
0.8
1
time (msec)
VC = 0 mV
m
n
h n4
m3h
Fig. 1.12 Response of the activation and inactivation variables m, h, and n to a step in voltage
46.
1.10 The ActionPotential Revisited 25
1.10 The Action Potential Revisited
In summary, the Hodgkin–Huxley model is a system of four differential equations;
there is one equation for the membrane potential and three equations for channel gat-
ing variables. In the case of a space-clamped squid axon, we write these equations as
cM
dV
dt
D N
gNam3
h.V ENa/ N
gKn4
.V EK/ N
gL.V EL/;
dn
dt
D Œ˛n.V /.1 n/ ˇn.V /n;
dm
dt
D Œ˛m.V /.1 m/ ˇm.V /m;
dh
dt
D Œ˛h.V /.1 h/ ˇh.V /h: (1.43)
Here, we added a parameter ; this is the temperature factor. It is important to real-
ize that the temperature at which an experiment is done can be very important. Since
channels are stochastic in nature, they are sensitive to the temperature, so the rates
of switching states depend exponentially on the temperature. Higher temperatures
cause faster switching. Thus, there is a factor
D Q.T Tbase/=10
10 : (1.44)
Q10 is the ratio of the rates for an increase in temperature of 10ı
C. For the squid
giant axon, Tbase D 6:3ı
C and Q10 D 3:
Figure 1.13 shows solutions of these equations in response to different levels
of steps in currents. Note that there is “all-or-none” behavior: When the applied
current is below some threshold, the membrane potential returns quickly to the rest;
when the current is above some threshold, there is an action potential. If the applied
current is sufficiently large and held for a sufficiently long time, then the model
generates a periodic response.
0 10 20 30 40
−60
−40
−20
0
20
40
time (msec)
V (mV)
0 50 100 150
−80
−60
−40
−20
0
20
40
time (msec)
V (mV)
Fig. 1.13 Responses of the Hodgkin–Huxley model to applied currents. Left transient responses
showing “all-or-none” behavior and right sustained periodic response
47.
26 1 TheHodgkin–Huxley Equations
−60
−20
20
0 2 4 6 8 10
time (msec)
VM
gNa
gK
Fig. 1.14 Solution of the Hodgkin–Huxley equations showing an action potential. Also shown are
the NaC
and KC
conductances
Figure 1.14 shows an action potential along with plots of the NaC
and KC
conductances, gNa and gK. Here, we start with the cell at rest and then depolar-
ize the cell by 10 mV at t D 0. The cell then generates a single action potential. In
Sect. 1.8, we described the events underlying the action potential in terms of the in-
ward and outward flow of NaC
and KC
ions. Here, we give a more “mathematical”
explanation in terms of the behavior of the dependent variables in the differential
equations.
When we depolarize the cell, we change the values of the activation curves:
n1.V / and m1.V / increase, whereas h1.V / decreases. Since n, m, and h tend
toward their activation curves, it follows that n and m initially increase, whereas h
decreases. That is, KC
channels open, whereas NaC
channels both activate and in-
activate. However, m is much smaller than both h and n. It follows that the NaC
channels activate much faster than they inactivate or KC
channels open. Therefore,
the NaC
conductance, gNa D N
gNam3
h, increases faster than gK D N
gKn4
.
The increase in the NaC
conductance leads to a large increase in the NaC
current,
INa D gNa.V ENa/. As long as the cell is near rest, the driving force V ENa
is large (recall that ENa C55 mV). Hence, the NaC
current will dominate the
equation for the membrane potential and V will increase toward the NaC
Nernst
potential. As V increases, m1.V / increases further, leading to further increase in
NaC
activation.
As V increases toward ENa, NaC
channels inactivate. This is because h !
h1.V / 0. Moreover, the NaC
driving force V ENa decreases. For both rea-
sons, the NaC
current turns off. Meanwhile, the KC
channel activates because
n ! n1.V / 1. Moreover, the KC
driving force V EK becomes very large.
48.
1.11 Bibliography 27
Vincreases
Na+ channels
activate: m
Na+ enters
cell
Na+ channels
inactivate: h
K+ channels
activate: n
Na+ deactivates: m
Na+ deinactivates: h
K+ deactivates: n
EK
ENa
Vrest
V
time
Fig. 1.15 Mechanisms underlying the action potential
It follows that eventually, the KC
current dominates and the membrane potential
must fall back toward the KC
Nernst potential. This corresponds to the downstroke
of the action potential.
After the action potential (Fig. 1.15), the cell is hyperpolarized with m1 0;
n1 0, and h1 1. After some time, m, n, and h approach their steady-state
values and the cell returns to rest.
1.11 Bibliography
There are many standard neuroscience textbooks that present the biological as-
pects covered in this chapter in much more detail. These textbooks include those
by Kandel et al. [144], Hille [122], and Martin [192]. The reader is also highly
recommended to look at Hodgkin and Huxley’s original papers [124]. A review
of these papers, along with a short review of the history leading up to them, is
given in Rinzel [229]. Excellent textbooks which emphasize modeling and quanti-
tative approaches are those of Johnston and Wu [139], Koch [156], Jack et al. [137],
Izhikevich [136], and Dayan and Abbott [54]. Keener and Sneyd [148] and Fall
et al. [83] give detailed introductions to mathematical aspects of cellular biophysics.
49.
28 1 TheHodgkin–Huxley Equations
1.12 Exercises
1. Suppose the external potassium in a mammalian cell is increased by a factor of
10. What is the new value of EK?
2. At 10ı
C a cell contains 80 mM sodium inside and has only 100 mM sodium
outside. What is the equilibrium potential for sodium?
3. Compute the resting potential for the mammalian cell using the same perme-
abilities as were used for the squid axon and the ion concentrations listed in
Table 1.1.
4. Derive the Nernst equation (1.2) from the Nernst–Planck equation by setting the
current to zero and integrating with respect to x across the membrane.
5. Compute the calcium equilibrium potential for a mammalian cell assuming that
the extracellular concentration is 5 mM and the intracellular concentration is
104
mM.
6. Complete the derivation the constant-field equation (1.3) from the linear Nernst–
Planck equation.
7. Derive the GHK equation (1.4) from the constant-field equation.
8. Consider the GHK equation and plot the I–V relation for different values of
the inside and outside concentrations. Show that for ŒCout ŒCin (respectively
ŒCout ŒCin) the I–V plot is concave down (respectively up).
9. Consider a passive, spherical cell with radius 0:003 cm2
, a resting membrane
potential of 65 mV, a membrane capacitance of 1F=cm2
, and a membrane
resistance of RM D 700 cm2
. Suppose the cell is injected with an applied
current of 5 nA/m2
for 2 s and then the current is turned off. What is the mem-
brane potential at t D 1; t D 2, and t D 3?
10. Suppose a passive axon has a diameter of 0.5 mm, a resting membrane resis-
tance of RM D 700 cm2
, and a transmembrane resistance of RL D 30 cm.
Compute the space constant. If the axon is 5 cm long and one end of the axon is
held at 120 mV above rest, then what is the potential at the other end?
11. (Johnston and Wu [139], page 12) The membrane capacitance of a typical cell
is 1 F=cm2
and the concentration of ions inside and outside the cell is about
0.5 M. Calculate the fraction of uncompensated ions on each side of the mem-
brane required to produce 100 mV in a spherical cell with a radius of 25 m.
12. Numerically solve the Hodgkin–Huxley equations. Start the system at rest and,
at some later time, inject an applied current to generate an action potential. Plot
the time courses of the NaC
and KC
conductances, as well as the gating vari-
ables m, h and n.
13. Numerically perform space-clamp experiments. That is, start the Hodgkin–
Huxley model at rest and, at some later time, change the membrane potential
and keep it as some “clamped” level. Plot the NaC
and KC
conductances for
when the membrane potential is stepped to different values.
50.
Chapter 2
Dendrites
We nowpresent mathematical theories for describing dendrites. Dendrites are very
important for many reasons. Indeed, the majority of the total membrane area of
many neurons is occupied by the dendritic tree. Dendrites enable neurons to connect
to thousands of other cells, far more than would be possible with just a soma, as
there is a huge membrane area to make connections. Dendrites may direct many
subthreshold postsynaptic potentials toward the soma, which summates these inputs
and determines if the neuron will fire an action potential. In addition to the treelike
structure of dendrites, many dendrites have additional fine structures at the ends
of the branches called spines. During development, animals that are raised in rich
sensory environments have more extensive dendritic trees and more spines.
Here, we will mainly discuss classical models for dendritic structure, activity,
and function. There has been tremendous progress recently in understanding the role
dendrites play in, for example, learning and neuronal computations, and the classical
view of dendrites is changing. Dendrites were originally thought, for example, to be
passive, in which the conductances and currents are constant; however, it is now
recognized that dendrites may have active voltage-gated channels and these may
greatly influence the firing properties of the neuron and how the neuron responds
to synaptic inputs. At the end of this chapter, we will briefly describe more recent
developments in understanding dendritic function.
2.1 Multiple Compartments
A very useful way to treat complicated dendritic structures is the compartmental ap-
proach. Here, one divides the dendritic tree into small segments or compartments
that are all linked together. Examples are shown in Fig. 2.1. Each compartment
is assumed to be isopotential and spatially uniform in its properties. Differences
in voltage and nonuniformity in membrane properties, including diameter, occur
between compartments rather than within them.
As a simple example, consider a two-compartment model in which each compart-
ment is viewed as an isopotential cylinder with radius ai and length Li . Let Vi be the
membrane potential of the ith compartment and let ci and rMi be the corresponding
G.B. Ermentrout and D.H. Terman, Mathematical Foundations of Neuroscience,
Interdisciplinary Applied Mathematics 35, DOI 10.1007/978-0-387-87708-2 2,
c
Springer Science+Business Media, LLC 2010
29
51.
30 2 Dendrites
Fig.2.1 (a) Branched
dendrite converted to a series
of cylinders for modeling.
(b) Simple
three-compartment model
a
b
2
3
1
specific membrane capacitance and specific membrane resistivity, respectively. We
assume each compartment has an electrode current and the total electrode current is
given by Ii
electrode. Finally, we assume the intracellular, or longitudinal, resistivity is
given by rL.
Now the capacitive and ionic currents for each compartment must be balanced
by the longitudinal and electrode currents. That is,
ii
cap C ii
ion D ii
long C ii
electrode; (2.1)
where ii
cap and ii
ion are the capacitive and ionic currents per unit area of membrane
for compartment i. As before,
ii
cap D ci
dVi
dt
and ii
ion D
Vi
rMi
(2.2)
if we assume the resting potential is 0. To compute ii
long, we need to determine the
total axial resistance. Note that the total resistance between the centers of the two
compartments is simply the sum of the two resistances of the half-cylinders that
separate the compartment centers. That is, the total resistance is given by
Rlong D
rLL1
2a2
1
C
rLL2
2a2
2
: (2.3)
Using Ohm’s law, we can write the expressions for the current from compartment i
to compartment j as
i1
long D g1;2.V2 V1/ and i2
long D g2;1.V1 V2/: (2.4)
52.
2.1 Multiple Compartments31
The coupling terms g1;2 and g2;1 are obtained by inverting (2.3) and dividing by the
surface area of the compartment of interest. That is,
g1;2 D
a1a2
2
rLL1.a2
2L1 C a2
1L2/
and
g2;1 D
a2a2
1
rLL1.a2
2L1 C a2
1L2/
:
Finally, to compute ii
electrode, we divide the total electrode currents by the surface
areas of the compartments. That is,
ii
electrode D
Ii
electrode
Ai
;
where Ai D 2ai Li is the surface area of compartment i.
Putting this all together, we find that the equations for two connected cylin-
ders are
c1
dV1
dt
C
V1
rM1
D g1;2.V2 V1/ C
I1
electrode
A1
;
c2
dV2
dt
C
V2
rM2
D g2;1.V1 V2/ C
I2
electrode
A2
: (2.5)
If instead of using conductances, gi;j , we use r1 D 1=g1;2 and r2 D 1=g2;1 then
we can express this system as
c1
dV1
dt
C
V1
rM1
D
V2 V1
r1
C i1;
c2
dV2
dt
C
V2
rM2
D
V1 V2
r2
C i2; (2.6)
where ii D Ii
electrode=Ai .
We can now explore the effects of two compartments on the input resistance
of the “cell.” Suppose we inject current into cell 1 only. Moreover, each cylinder
is identical, with the same length and radius. Then, r1 D r2 r. What is the
input resistance due to the coupling? To find this, we must compute the steady-state
potential due to the coupling. Without loss in generality, define rM D rM1 D rM2:
A simple bit of algebra shows that
V1=i1 D
rM.r C rM/
r C 2rM
53.
32 2 Dendrites
andthus the ratio of the coupled to the uncoupled input resistance is
Rinput;coupled
Rinput;uncoupled
D 1
rM
r C 2rM
I
that is, the input resistance decreases. To get the same increment in potential, the
current required for the coupled system is more than that required for the uncoupled
system because some current is drained away by the second compartment.
In a similar way, we can derive a compartmental model for a general treelike
structure. A general algorithm for computing the correct equations is
For each cylinder, j, with radius and length aj and Lj in micrometers,
compute the surface area, Aj D 2aj Lj , and the axial resistance factor,
Qj D Lj =.a2
j /:
The membrane capacitance is Cj D cj Aj 108
and the membrane resistance
is Rj D .rMj =Aj / 108
.
The coupling resistance between compartments j and k is Rjk D rL
2
.Qj C
Qk/ 104
.
The equations are then
Cj
dVj
dt
D
Vj
Rj
C
X
k connected j
Vk Vj
Rjk
C Ij :
The factors of 10˙8
and 104
are the conversion from micrometers to centimeters.
For example, consider a two-compartment model with (1) compartment 1 having a
length of 200 m and radius of 30 m and (2) compartment 2 having a length of
20 m and radius of 20 m. Then, R1 D 2:65 107
; C1 D 3:77 1010
F,
R2 D 3:98 108
; C2 D 2:52 1011
F, and Rlong D 4:34 104
: Thus,
10
dV1
dt
D V1 C 611.V2 V1/; 10
dV2
dt
D V2 C 9;181.V2 V1/;
where time is in milliseconds and the coupling coefficients are dimensionless. Note
how the ratio of the coupling strengths is the same as the reciprocal of the area ratios.
The bigger compartment has a much greater effect on the smaller compartment than
vice versa.
We also remark that the standard units used in most compartmental models are
microfarads per square centimeter, millisiemens per square centimeter, and mi-
croamperes per square centimeter for the capacitance, conductance, and applied
current, respectively. Experimentalists do not generally know the current density,
but only the total current injected. Typical currents injected into a cell are of the
order of less than 1 nA.
To generate arbitrary compartmental models, one needs only to compute the
length, diameter, and the connectivity of the cylinders that make up the dendritic
tree. The program NEURON [33] enables an experimentalist to input a digitized
picture of a neuron and the program will automatically produce a compartmental
model of the neuron by linking together many cylinders.
54.
2.2 The CableEquation 33
2.2 The Cable Equation
Mathematically, dendrites and axons are regarded as continuous media rather than a
series of compartments. Previously, we derived the cable equation for a simple cable
in which the radius along the cable was assumed to be constant. Here we derive the
cable equation for more general geometries. This is done by considering the limit as
the number of compartments in an approximation of it tends to infinity.
Suppose the cable is defined on the interval .0; `/ with a circular cross-section
and diameter d.x/. We break the cable into n pieces and define xj D jh where
h D `=n: Each piece has a surface area Aj D dj h where dj D d.xj /, and
crossectional area, d2
j =4: Let cM and rM denote the specific membrane capacitance
and resistance, and let rL be the longitudinal resistance. Then, neglecting the end
points, the voltage satisfies:
cMAj
dVj
dt
D
Vj
rM=Aj
C
VjC1 Vj
4rLh=.d2
j C1/
C
Vj 1 Vj
4rLh=.d2
j /
:
Note that we use the larger diameter for the transmembrane resistance; in simula-
tions, the average of the two would be preferred. Dividing by h the coupling term
simplifies to:
h
d2
j C1.VjC1 Vj /
4rLh
d2
j .Vj Vj 1/
4rLh
!
:
As h ! 0, this goes to the diffusion operator:
4rL
@
@x
d2
.x/
@V
@x
:
Thus, (dividing by d.x/ the cable equation has the form:
cM
@V
@t
D
V
rM
C
1
4rLd.x/
@
@x
d2
.x/
@V
@x
: (2.7)
We remark that the term
d2
j .Vj1 Vj /
4rLh
has dimensions of current and in the limit as h ! 0 is called the longitudinal
current:
IL D
d2
.x/
4rL
@V
@x
: (2.8)
If one is interested only in the passive cable and d.x/ D d is constant, then it is
convenient to multiply both sides by rM obtaining the linear cable equation:
@V
@t
D V C 2 @2
V
@x2
(2.9)
the steps leadingup to an orchestra, the fringe of her mask confined
by her hand, so as to prohibit even a glimpse of her ivory chin, she
subjected to a rigid scrutiny all who issued from the conservatory.
Suddenly, from the door nearest to her, the hussar and his
companion made their appearance, and, as they passed, she
shrouded herself behind the portly figure and sumptuous
embroideries of a Venetian doge. Then she resumed her watch, and
a minute had not elapsed when she saw the tall black domino,
whom she had observed during the evening, re-enter the dancing-
room and make his way as fast as the crowd would allow him to the
nearest door of exit, with a hurried and irregular step, hardly to be
explained otherwise than by sudden illness or violent emotion. She
followed him to the head of the staircase, down which he rushed,
disappearing at its foot through the crowd of lackeys in the hall.
Having seen this, she re-entered the ball-room, sought out the
hussar and his companion, and soon afterwards was whirling with
the former in the giddy circles of a waltz.
Some hours later, as the Hungarian retired from the ball, almost
borne along in the dense stream of masks that now flowed through
the rooms, he felt a momentary pressure of his hand. A paper
remained in its palm, upon which his fingers mechanically closed.
Amidst the ever-moving throng it was impossible to detect the
person from whom he had received it. By this time a large portion of
the company, oppressed by the heat, had unmasked, but he knew
none of the faces he saw around him, whilst of those who had
preserved their vizards he could fix on none as object of suspicion.
So soon as he could extricate himself from the crowd, he unfolded
the paper. It contained the following mysterious words, hastily
scrawled with a pencil:—
One whom you think asleep wakes and watches. He is here; has
followed and overheard you, and will seek revenge. Be prepared.
Proof is difficult: denial may be safety. Adopt it at all risks. Masked,
the sisters are undistinguishable. Credit this warning from a sincere
friend.
57.
Thrice the Hungarianperused this mysterious billet; and then,
thrusting it into the breast of his richly braided jacket, slowly left the
house.
CHAP. III.—THE ACCUSATION.
The house selected by Baron Ernest von Steinfeld, wherein to pass
what might possibly be his last season in Paris, was situated in the
Rue St Lazare. It was one of those buildings, of frequent occurrence
in modern Parisian architecture, which seem intended to gratify the
taste of such persons as prefer the English fashion of occupying an
entire house, to the French one of dwelling upon a floor. At the
bottom of a paved court-yard, around three sides of which was built
a large mansion containing many tenants, stood one of those
edifices known in French parlance as pavilions—not that they
possess a dome, resemble a tent, or, for the most part, have any of
the qualities of a summer-house, but because, in Paris, the term
house is grudgingly bestowed upon a building of less than five
stories and thirty or forty rooms. This pavilion had but three stories
and a dozen rooms; it was a particularly complete and independent
habitation, standing well back from the body of the house, under
whose number it was included, and of which, although detached, it
was considered to form part; and having two entrances, one through
the court, the other from a lane running at right angles with the
street. The ground-floor contained, besides a light and commodious
vestibule and servant's offices, only one apartment, a handsome
dining-room, in which, however, it was impossible, for three quarters
of the year, to dine without lamps—the daylight admitted by its one
broad window being greatly limited by the walls of a nook of garden,
and by the impending branches of a laburnum and acacia, which
mingled their boughs in affectionate union, twin lords of a square
yard of grass, and of a fathom's length of flower-bed, and in the
spring-time rejoiced the inmates of the pavilion with the odorous
rustle of their yellow clusters and rose-coloured blossoms. The first
58.
floor contained twopleasant drawing-rooms and a boudoir; the
second, bath, bed, and dressing rooms. The roof, flat and
surrounded by a parapet, commanded a view over the adjacent
gardens of an extensive bathing establishment and maison de santé,
and was no unpleasant resort, on a fine day, for persons desirous to
inhale the fresh air, or to scent it with the fumes of Havana's weed.
This pavilion, described by the Petites Affiches as fraîchement décoré
—the said decoration consisting in fresh paint and paper, and in a
profusion of that cheerful French luxury, large and excellent mirrors
—was rented for six months by Baron Steinfeld, who had hired, for
the same period, from a fashionable upholsterer—for a sum which
would almost have furnished the house permanently in a plainer
manner—a complete set of furniture, against whose perfect elegance
and good taste not a syllable could be breathed. His establishment
was as correct as his residence. It consisted, in the first place, of a
French cook, with whose sauces Arthur de Mellay had repeatedly
expressed his willingness to eat a fragment of his father; which offer
—considering the worthy count had been a guardsman in the time of
Louis XVI., and, consequently, was neither young nor tender—was
certainly a high testimonial to the merits of sauce and cook. Then
came an Italian valet, quite as skilful a personage in his way as the
professor of gastronomic science—speaking three or four languages,
accumulating in his own individuality the knowledge and
acquirements of a legion of hairdressers, tailors, perfumers, and the
like—thoroughly versed in the arcana of the toilet, a secretary in
case of need, and a perfect Mercury in matters of intrigue. The third
person of Steinfeld's household, the last, and also by much the least
—physically speaking, that is to say, but by no means in his own
estimation—was one of those miniature tigers, (copied from the
English, and essential appendages to the establishment of a Paris
lion,) who look as if they had been subjected to that curious Chinese
process by which lofty shrubs and forest trees are stunted to
dimensions that permit the plantation of a grove in a flower-pot—
wizen-faced, top-booted abortions, uniting the mischief and the
proportions of a monkey, and frightfully precocious in every species
of villany. The house also contained, during the day, an old
59.
Frenchwoman, of aspecies indigenous and confined to Paris—the
patient butt of the cook's ill-humours and of the groom's pranks,
with bearded chin and slipshod feet, and willing for any sort of dirty
work, from the scouring of a kettle to the administration of the
remedy renowned in French pharmacy.
It was an hour past noon on the day succeeding the Countess of M
——'s masquerade, and Steinfeld sat alone at breakfast. It were
more correct to say that he sat at the breakfast table; for the
savoury meal before him was still untasted, and he seemed in no
haste to attack it. In vain the green oysters from Ostend lay
invitingly open, and one of Chevet's pies displayed, through a
triangular aperture in its crust, the tender tints of an exquisite foie-
gras—the result of the martyrdom of some unhappy Strasburg duck;
in vain a fragrant steam of truffles oozed from beneath the covers of
two silver dishes, fresh from the laboratory of Macedoine the cook,
and mingled its odours with the flowery aroma of a bottle of
Sauterne, from which Rufini the valet had just extracted the long
yellow-sealed cork. Apparently, none of these creature-comforts
dwelt in the desires of the baron, who sat sideways to the table, his
chin resting on his hand, gazing upon vacancy with an intenseness
bespeaking deep preoccupation. One acquainted with Steinfeld's
circumstances would have hesitated little in conjecturing the nature
of the unpleasant reflections in which he seemed absorbed. They
might very well have for motive the unprosperous state of his
exchequer, the heavy incumbrances weighing upon the hereditary
acres, the approaching decease of that convenient but fickle ally, on
whose succour half the world exist, and whose name is Credit. The
baron had been any thing but a prudent man. Too careless of the
future, he had neglected fortune when she offered herself to his
embrace; and now she revenged herself by averting her
countenance. Of high descent and fair estate, handsome person and
fascinating manners, for some years Steinfeld might have aspired to
the hand of almost any heiress in Vienna or Paris. Numerous were
the matrimonial overtures that had been more or less directly made
to him, at a time when, in love with his bachelorhood, and
60.
celebrated for hisbonnes fortunes, he looked upon the bonds of
Hymen as the most oppressive of fetters, intolerable even when
sheathed in gold. The matchmakers, repulsed without exception, at
last renounced all further attempts upon the hand of the handsome
Austrian—as Steinfeld was generally called in Paris—and declared
him an incorrigible partisan of celibacy. To the unmolested
enjoyment of his bachelor bliss the baron was for some years left,
until one morning he awoke to the disagreeable consciousness that
profuse expenditure had done its work, and that ruin or a rich
marriage were the only alternatives left him. He was fully alive to the
difficulties placed in the way of the latter by the change in his
circumstances. His ancient name and personal advantages remained,
but his fair estate was in the hands of the harpies; and however
disposed romantic young ladies might be to overlook this misfortune,
prudent papas would deem it a serious stumbling-block. Then it was
that, roused by horrid visions of approaching poverty from his usual
state of happy insouciance, the baron gathered together the relics of
his past opulence, squeezed and exhausted every remaining
resource, and, assuming a bold front against bad fortune, returned
to Paris, with much the feelings of the soldier who screws up all his
energies to conquer or to die. It was no apprehension, however, as
to the result of this final struggle—no nervous trepidation arising
from the imminence of his situation, that now clouded Steinfeld's
brow and spoiled his appetite. On the contrary, he deemed victory
secure, and beheld himself, in no remote perspective, emerging
triumphantly from his difficulties, even as a snake, casting its shabby
skin, reappears in glittering scales of gold. He had not wasted the
three months he had passed in Paris, and was well satisfied with the
result of his exertions. His present uneasiness had a different origin
—one similar to the cause by which, some fifteen hours previously,
we saw Sigismund Fatello so deeply moved. The baron turned and
twisted in his hand a letter, to whose contents he again and again
recurred, pondering them intently. Like that received by the banker,
the billet was anonymous; like his, it contained but three or four
lines; but, despite its brevity and want of authenticity, it proved, on
the part of the writer, whoever that might be, an acquaintance with
61.
the baron's mostimportant secret, that did not fail greatly to
disquiet him. Who had thus detected what he deemed so surely
concealed? He strained his eyes and memory, in vain endeavouring
to recognise the handwriting; and, more than once, fancying he had
done so, he fetched notes and letters from a desk in the adjoining
boudoir, to compare them with the anonymous epistle. But the
comparison always dissipated his suspicion. Then, taking a pen, and
a diminutive sheet of amber-scented paper, he began a note, but
tore the paper after writing only three words, and threw the
fragments impatiently into the fire. Just then the pavilion bell rang
loudly; the next minute there was a knock at the room door, and
Celestin the tiger made his appearance, bearing a card inscribed
with the name of M. Sigismund Fatello, and an inquiry whether
Monsieur le Baron was at home and visible.
On reading the banker's name, Steinfeld made a slight and sudden
movement, almost amounting to a start, but, instantly recovering
himself, he bade his groom show the visitor up stairs. At the same
time he hastily seated himself, ordered Rufini to take off the covers,
poured some wine into a glass, and helped himself from the first
dish that came to hand; so that when Fatello, ushered in by the
groom, entered the apartment, he had all the appearance of one
whose whole faculties were concentrated, for the time being, in the
enjoyment of an excellent meal. Rising from his chair, with an air of
jovial cordiality, he hastened to welcome the banker.
An unexpected pleasure, my dear Fatello, said he. What
favourable chance procures me so early a visit? You are come to
breakfast, I hope. Rufini, a knife and fork for M. Fatello.
I have breakfasted, M. le Baron, replied Fatello, with a dryness
amounting almost to incivility. If my call is untimely, my business is
pressing——and private, he added, with a glance at the Italian, who
stood in respectful immobility behind his master's chair.
Leave the room, Rufini, said Steinfeld.
62.
The well-drilled valetbowed in silence, and glided noiselessly from
the apartment.
Now then, my good friend, said the Austrian, in the same gay
offhand tone as before, I am all ear and attention. What is up?
Nothing bad, I hope; nothing so serious as to spoil my appetite. I
have heard a proverb condemning discourse between a full man and
a hungry one.
Fatello made no immediate reply. There was something very peculiar
in his aspect. His lips were pale and compressed, and his brows
slightly knit. He seemed constraining himself to silence until he felt
he could speak calmly on a subject which roused anger and
indignation in his breast. Whilst seemingly engrossed by his
breakfast, Steinfeld lost not a look or motion of his visitor's, not a
line of his physiognomy, or a glance of his small piercing eye. And
the baron, notwithstanding his assumed careless levity of manner,
did not feel altogether at his ease.
You have not turned conspirator, I hope, said he, when Fatello,
after a short but awkward pause, still remained silent. No Henri-
quinquist plot, or plan to restore the glorious days of the guillotine
and the Goddess of Liberty? No, no; a Crœus of your calibre, my
dear Fatello, would not mix in such matters. Your plotters are hungry
dogs, with more debts than ducats. Talking of hunger—I am grieved
you have breakfasted. This mushroom omelet does honour to
Macedoine.
The baron would have talked on,—for at that moment any sort of
babble seemed to him preferable to silence. But Fatello, who had not
heard a word he had said, suddenly rose from his seat, rested his
hands upon the table, and leaning forward, with eyes sternly fixed
upon Steinfeld, uttered these remarkable words, in tones rendered
harsh and grating by the effort that made them calm:
Monsieur le Baron de Steinfeld, you are courting my wife!
63.
The most expertphysiognomist would have failed to detect upon the
countenance of the ex-diplomatist any other expression than one of
profound astonishment, tinged by that glow of indignation an
innocent man would be likely to feel at an unfounded accusation,
abruptly and brutally brought. After sustaining for a few seconds
Fatello's fixed and angry gaze, his features relaxed into a slightly
contemptuous smile.
The jest is surely in questionable taste, my dear M. Fatello. And the
severity of your countenance might alarm a man with a conscience
less clear than mine.
I jest not, sir, with my honour and happiness, retorted Fatello, with
a rude fierceness that brought a flush to the baron's cheek—a flame
of anger which the next moment, however, dispelled.
Then, my dear M. Fatello, said Steinfeld, since, instead of a bad
jest, you mean sober earnest, I can only say you are grossly
misinformed, and that your suspicions are as injurious to Madame
Fatello, as your manner of expressing them is insulting to myself.
I have no suspicions, replied Fatello, but a certainty.
Impossible! said the baron. Name my accuser. He shall account
for the base calumny.
He desires no better, replied Fatello, sternly. I myself accuse you.
No slanderous tongues, but my own ears, are evidence against you.
And yourself, sir, shall confess what you now so stubbornly deny. You
were at last night's masquerade.
I was so.
In hussar uniform—crimson vest and white pelisse.
Steinfeld bowed assent. The uniform of the regiment to which I
formerly belonged.
A black domino was on your arm.
64.
Ma foi! criedthe baron, with a laugh that sounded rather forced,
if you demand an account of all the masks I walked and danced
with, I shall hardly be able to satisfy you. Dominos there were,
doubtless; and, of all colours, black amongst the rest.
You equivocate, sir, said Fatello, angrily. I will aid your memory.
The domino I mean was your companion early in the night. The
domino I mean danced once with you, (a waltz,) and afterwards
walked with you through the rooms, in deep conversation. The
domino I mean stood with you for more than ten minutes beside the
fountain in the conservatory. The domino I mean was my wife; and
you, Baron Steinfeld, are a villain!
During this singular conversation Steinfeld had sat, leaning back in
his large elbow-chair, in an attitude of easy indifference—one
slippered foot thrown carelessly over the other, and his hands thrust
into the pockets of his damask dressing-gown. On receiving this last
outrageous insult, his lip blanched with passion, his whole person
quivered as with an electric shock, and he half rose from his semi-
recumbent position. But the baron was a man of vast self-command;
one of those cool-headed cool-hearted egotists who rarely act upon
impulse, or compromise their interests by ill-timed impetuosity. The
first choleric movement, prompting him to throw Fatello down stairs,
was checked with wonderful promptitude, and with little appearance
of effort. In reality, however, the effort was a violent one. As a
soldier at the triangles bites a bullet with the rage of pain, so
Steinfeld clenched his hands till the strong sharp nails almost cut
into the palm. As he did so, a paper in his pocket rustled against his
knuckles. It was the note so mysteriously conveyed to him at the
masquerade, and which he had been pondering when Fatello was
announced. To one so quick-witted, the mere touch of the paper was
as suggestive as a volume of sage counsels. In an instant every sign
of annoyance disappeared from his features; he rose quietly from his
seat, and with easy dignity and an urbane countenance, confronted
Fatello, who stood gloomy and lowering before the fire.
65.
I see, M.Fatello, he said, that you are bent upon our cutting each
other's throats; but, strange as it may seem, after the terms you
have employed, I still hope to avert the unpleasant necessity. For
one moment moderate your language, and give me time for brief
explanation. If I rightly understand you, it is from your own
observations you thus accuse me; and I presume you did me the
honour of a personal surveillance at last night's ball?
Fatello, his violence checked for the moment from further outbreak
by the baron's courtesy and coolness, made a gesture of sullen
assent.
And that you overheard a part, but not the whole, of my
conversation with the black domino in question?
I heard enough, and too much, replied Fatello, with a savage scowl
at his interlocutor. This is idle talk, mere gain of time. Baron
Steinfeld! cried the banker, in a voice that again rose high above its
usual pitch, you are——
Stop! interrupted Steinfeld, speaking very quickly, but with an
extraordinary and commanding calmness, which again had its effect.
Descend not to invective, M. Fatello. There is always time for
violence. Hear reason. You are in error, an error easily explained. I
certainly saw Madame Fatello at the ball, saw and spoke with her—
patience, sir, and hear me! But the domino, of my conversation with
whom you heard a part, was not Madame Fatello, but Mademoiselle
Gonfalon. You take little interest in the frivolities of a masquerade,
and are possibly unaware that the two ladies' dresses were exactly
similar. You can have heard our conversation but imperfectly, or you
would not have wronged me by this suspicion.
Whilst uttering these last sentences, Steinfeld redoubled the
keenness of the scrutiny with which he regarded the banker's
uncomely and agitated physiognomy. But although piquing himself,
as a former diplomatist, on skill in reading men's thoughts through
their faces, he was unable to decipher the expression of Fatello's
66.
countenance on receivingthis plausible explanation of the error into
which he had been led by the sisters' identity of costume. As he
proceeded with it, the banker's lips, slightly parting, gave his face an
air of stupefied wonderment, in addition to its previously inflamed
and angry aspect. When Steinfeld concluded an explanation uttered
with every appearance of sincerity and candour, and in that flexible
and affable tone which, when he chose to employ it, imparted to his
words a peculiarly seductive and persuasive charm, Fatello's lips
were again firmly closed, and curled with a curious and inexplicable
smile. This faded away; he struck his left hand against his forehead,
and remained for some moments plunged in thought, as if he hastily
retraced in his memory what he had heard the night before, to see
how it tallied with the explanation just given him. Thus, at least,
Steinfeld interpreted his manner; and although the Austrian's
countenance preserved its serenity, his heart throbbed violently
against his ribs during the banker's brief cogitation. The result of this
was evidently satisfactory to Fatello, from whose brow, when his
hand again dropped by his side, the lowering cloud had disappeared,
replaced by affability and regret.
I see, he said, with better grace than might have been expected
from him, and taking a step towards Steinfeld, that nothing remains
for me but to implore your pardon, baron, for my unwarrantable
suspicions, and for the harsh and unbecoming expressions into
which they betrayed me. Jealousy is an evil counsellor, and blinds to
the simplest truths. I scarce dare hope you will forgive my
intemperate conduct, without exacting the hostile meeting for which
I was just now as eager as I at present am to avoid it. If you insist, I
must not refuse, but I give you my word that if I have a duel with
you to-day, nothing shall induce me to depart from the defensive.
I should be unreasonable, replied Steinfeld graciously, if I exacted
ampler satisfaction than this handsome apology, for what, after all,
was no unnatural misconception. Ten years ago, I might have been
more punctilious, but after three or four encounters of the kind, a
67.
duel avoided, whenits real motive is removed, is a credit to a man's
good sense, and no slur upon his courage.
No one will ever attack yours, my dear baron, said Fatello. I only
hope you will always keep what has passed between us this morning
as profound a secret as I, for my own sake, certainly shall do. I am
by no means disposed to boast of my part in the affair.
Steinfeld bowed politely, and the two men exchanged, with smiles
upon their faces, a cordial grasp of the hand.
Out of evil cometh good, said the banker sententiously, subsiding
upon the silken cushions of a causeuse that extended its arms
invitingly at the chimney-corner. I am delighted to find that the
leaden bullet I anticipated exchanging with you is likely to be
converted into a golden ring, establishing so near a connexion
between us as to render our fighting a duel one of the least
probable things in the world. My dear baron, I shall rejoice to call
you brother-in-law.
It would be a great honour for me, replied Steinfeld, but you
over-rate the probability of my enjoying it. Nothing has passed
between Mademoiselle Gonfalon and myself to warrant my reckoning
on her preference.
Tush, tush! baron, said Fatello, apparently not heeding, or not
noticing the somewhat supercilious turn of Steinfeld's phrases, you
forget the new and not very creditable occupation to which the
demons of jealousy and suspicion last night condemned me. You
forget that I tracked you in the promenade, and lay in ambush by
the fountain, or you would hardly put me off with such tales as
these.
The baron winced imperceptibly on being thus reminded how closely
his movements had been watched.
You are evidently new at the profession of a scout, said he
jestingly, or you would have caught more correctly my conversation
68.
with your amiablesister-in-law. Mademoiselle Gonfalon is a charming
person; the mask gives a certain license to flirtation, and a partial
hearing of what passed between us has evidently misled you as to
its precise import.
Not a bit of it! cried Fatello, with an odd laugh—I heard better
than you think, I assure you; and what I did hear quite satisfied me
that you are a smitten man, and that Sebastiana is well disposed to
favour your suit.
I must again protest, said Steinfeld, expressing himself with some
embarrassment, that the thought of becoming Mademoiselle
Gonfalon's husband, great as the honour would be, has never yet
been seriously entertained by me; and that, however you may have
been misled by the snatches of our conversation you overheard,
nothing ever passed between us exceeding the limits of allowable
flirtation—the not unnatural consequence of Mademoiselle
Sebastiana's fascinating vivacity, and of the agreeable footing of
intimacy on which, for the last three months, I have found
admittance at your hospitable house.
Sigismund Fatello preserved, whilst the baron waded through the
intricacies of his artificial and complicated denial, a half-smile of
polite but total incredulity.
My dear baron, said he, gravely, when Steinfeld at last paused, I
am sure you are too honourable a man to trifle with the affections of
any woman. I know you as the very opposite character to those
heartless and despicable male coquets, who ensnare susceptible
hearts for the cruel pleasure of bruising or breaking them, and
sacrifice, in their vile egotism, the happiness of others to the
indulgence of paltry vanity. I detect the motives of your present
reserve, and, believe me, I appreciate their delicacy. Rumour, that
eternal and impertinent gossip, has asserted that Baron Ernest von
Steinfeld has impaired, by his open hand and pursuit of pleasure, the
heritage of his forefathers. I do not mean that this has become
matter of common report; but we bankers have opportunities of
69.
knowing many things,and can often read in our bill-books and
ledgers the histories of families and individuals. In short, it is little
matter how I know that your affairs, my dear baron, are less
flourishing than they might be, or than you could wish. But this,
after all, is an unimportant matter. The dirty acres are still there—the
Schloss Steinfeld still stands firm upon its foundation, and though
there be a bit of a mortgage on the domain, and some trouble with
refractory Jews, it is nothing, I am sure, but what a clear head, and
a little ready cash, will easily dispose of.
It was natural to suppose that a lover, whose position on the brink of
ruin made him scruple to ask the hand of his mistress of her nearest
male relative and protector, and who found his embarrassments
suddenly smoothed over and made light of by the very person who
might be expected to exaggerate them, would be the last man to
place fresh stumbling-blocks on the path to happiness thus
unexpectedly cleared before him. Steinfeld, however, appeared little
disposed to chime in with the banker's emollient view of his
disastrous financial position. With an eagerness that bespoke either
the most honourable punctiliousness, or very little anxiety to become
the husband of Mademoiselle Gonfalon, he set Fatello right.
I heartily wish, said he, matters were no worse than you suppose.
You quite underrate my real embarrassments. My estate is mine only
nominally; not a farthing it produces comes into my pocket; the very
castle and its furniture are pledged; some houses in Vienna, and a
few thousand florins of Austrian rentes, derived from my mother,
melted away years ago; I am deeply in debt, and harassed on all
sides by duns and extortioners. I calculated my liabilities the other
day—why, I know not, for I have no chance of clearing them—and I
found it would require three hundred thousand florins to release my
lands and pay my debts. You see, my dear M. Fatello, I am not a
very likely match for an heiress.
Fatello had listened with profound attention to the insolvent balance-
sheet exhibited by the baron.
70.
Three hundred thousandflorins—six hundred thousand francs, said
he, musingly—allowing for usury and overcharges, might doubtless
be got rid of for a hundred thousand less. Well, baron, when
Sebastiana marries, she will have more than that tacked to her
apron. Her father left her something like half a million, and I have
not let the money lie idle. She is a richer woman, by some thousand
louis d'ors, than she was at his death. I don't carry her account in
my head, but I daresay her fortune would clear your lands, and
leave a nice nest-egg besides. And although she certainly might find
a husband in better plight as regards money matters, yet, as you are
so much attached to each other, and happiness, after all, is before
gold, I shall make no difficulties. I noticed the girl was absent and
sentimental of late, but never guessed the real cause. Ah, baron!
you fascinating dogs have much to answer for!
Whilst Fatello thus ran on, with, as usual, more bluntness than good
breeding, Steinfeld was evidently on thorns; and at the first
appearance of a pause in the banker's discourse, he impatiently
struck in.
I must beg your attention, M. Fatello, said he, whilst I repeat
what you evidently have imperfectly understood—that it has never
entered my head to gain Mademoiselle Gonfalon's affections, and
that I have no reason to believe I should succeed in the attempt. I
again repeat that nothing but the most innocent and unimportant
flirtation has passed between us. I am deeply sensible of your kind
intentions—grateful for your generous willingness to overlook my
unfortunate circumstances, and to promote my marriage with your
sister-in-law; but, flattering and advantageous as such a union
would be to me, I am not certain it would lead to that happiness
which you justly deem preferable to wealth. I doubt whether my
disposition and that of Mademoiselle Sebastiana would exactly
harmonise. Moreover, necessitous though I am, it goes against my
pride to owe every thing to my wife. It would pain me to see her
dowry swallowed up by my debts. Let us drop the subject, I entreat
you. To-morrow you will appreciate and rejoice at my hesitation. I
71.
fully comprehend thegenerous impulse that prompts you. Having
done me an injustice, you would compensate me beyond my merits.
Thanks, my good friend; but, believe me, if happiness resides not in
wealth, neither is it found in hasty or ill-assorted unions. And, to tell
you the truth, however politic a rich marriage might be in the
present critical state of my affairs, I long ago made a vow against
matrimony, which I still hesitate to break.
You are the best judge of your own motives, said Fatello, stiffly,
but you quite misconstrue mine. It never entered my head to view
you as a victim, or to think myself called upon to atone, by providing
you with a rich and handsome wife, for the jealousy you so
successfully proved groundless. Such compensation would be
excessive for so slight an injury. No, no, baron—you have quite
mistaken me. As the nearest connexion and natural guardian of
Mademoiselle Gonfalon, it is my duty to watch over her, and not to
allow her feelings to be trifled with. For some time past, I have
suspected her affections were engaged, but it never occurred to me
they were fixed upon you. Well—last night I go to a ball, and,
actuated by suspicions to which it is unnecessary to recur, I listen to
your conversation with my sister-in-law. To a plain man like myself, it
bore but one interpretation—that you have sought and won her
heart. You deny this, and assert your language to have been that of
common gallantry and compliment, such as may be addressed to
any woman without her inferring serious intentions. Here, then, we
are gravely at issue. You maintain my ears deceived me; I persist in
crediting their evidence. Fortunately, an arbiter is easily found. I
shall now return home, see my sister-in-law, and confess to her my
eaves-dropping, keeping its real motive and my visit to you
profoundly secret. From her I shall learn how matters really stand. If
her account agree with Baron Steinfeld's, I shall evermore mistrust
my hearing; if the contrary, and that the baron, himself a sworn foe
to marriage, has compromised the happiness of a young and
confiding woman, why, then, he will not be surprised if I seek of
him, for so grave an offence, the reparation which a short time ago I
was ready to afford him for one comparatively insignificant. And
72.
Fatello bowed formally,and with severe countenance moved towards
the door. But before he could leave the room, Steinfeld, who had
stood for a moment thoughtful and perplexed, hurried to intercept
him, and laid his hand upon the lock.
You are really too hasty, Fatello, said he, and not altogether
reasonable. What ill weed have you trodden upon, that makes you
so captious this morning? Own that our conversation has taken an
odd turn! Would any one believe that you, Fatello the millionnaire,
press a marriage between your sister, the wealthy Mademoiselle
Gonfalon, and myself, the needy Baron Steinfeld—and that it is I, the
ruined spendthrift, from whom the obstacles to the match proceed?
Neither in romance nor in real life has the case a precedent. And you
may be assured the world will not applaud your wisdom, nor
Mademoiselle Sebastiana feel grateful for your zeal.
For the world's applause I care not that, replied Fatello, snapping
his fingers. As to my sister, I have neither will nor power to
constrain her. I do but afford her the protection she is entitled to at
my hands. I press her upon no man, but neither do I suffer her to
be trifled with. Sebastiana Gonfalon does not lack suitors, I can
assure you.
Unquestionably, said Steinfeld, with an absent air; Mademoiselle
Gonfalon is indeed a most charming person, and, were she
penniless, would still be a prize to any man. I only wish I enjoyed
the place in her good opinion you so erroneously imagine me to
occupy.
Well, well, said Fatello, striving to get at the door, before which the
baron had planted himself, since error there is, it will soon be
cleared up. You cannot blame me, baron, for preferring, in so
delicate an affair, the testimony of my own ears to that of any one
person. But if two unite against me, I shall think myself crazed or
bewitched, and shall at least be silenced and confounded, if not
entirely convinced.
73.
Answer me onequestion, said Steinfeld. If yesterday, before you
overheard a part of my conversation with your sister, I had asked of
you her hand, exposing to you at the same time the state of my
fortunes, or rather of my misfortunes, would you then have
sanctioned my suit and pleaded my cause with Mademoiselle
Gonfalon? Would you, and will you now—for, believe me, I need it
more than you think—add the weight of your arguments and
advocacy to the prepossession you persist in thinking your sister has
in my favour, a prepossession of whose existence I hardly dare
flatter myself?
Why not? said Fatello, with an air of straightforward cordiality.
Why not? You are not rich, certainly but Sebastiana is rich enough
for both. You have high birth, talents, interest with the Emperor,
and, once married, with your debts paid, and your wild oats sown,
you may take ambition instead of pleasure for a mistress, and aspire
to high employment. Why not return to diplomacy, for which you are
so admirably qualified, and come back to us as Austrian
ambassador? Believe me, baron, there is a fine career before you, if
you will but pursue it.
Perhaps, said Steinfeld, smiling to himself, like a man to whom a
bright perspective is suddenly thrown open; and, as you say, the
first step would be a suitable marriage, which, by ridding me of all
encumbrance, might enable me to climb lightly and steadily the hill
of wealth and honours.
And a millionnaire brother-in-law to give you an occasional push by
the way, added Fatello, with one of his heavy, purse-proud smiles;
pushes you may repay in kind, for diplomatist and financier should
ever hunt in couples.
My dear Fatello, said Steinfeld, the prospect is too charming to be
lightly relinquished. You must think strangely of my first reluctance
to avail myself of your friendly disposition in my favour; but I so little
suspected it, I was so bewildered by its sudden revelation, so
embarrassed by my own difficulties—and then pride, you know—a
74.
morbid fear ofbeing thought mercenary; in short, you will make
allowance for my strange way of meeting your kind encouragement.
I can only say, that since you deem me worthy of her, and if you can
obtain her consent, (a more difficult task, I fear, than you imagine,) I
shall be the happiest of men as the husband of the adorable
Sebastiana.
That is speaking to the purpose, said Fatello; and, for my part, I
repeat that I shall be happy to call you brother-in-law. I will do my
best for you with Sebastiana, to whom I will at once communicate
your formal demand in marriage. But, pshaw! you rogue, added he,
with a clumsy attempt at archness, you have made pretty sure of
her consent, and need no brotherly advocate.
Indeed you are mistaken, replied Steinfeld earnestly. I only wish I
were as confident, and with good reason, as you think me.
Well, well, no matter, said the banker. You shall shortly hear your
fate.
I shall be on thorns till I learn it, said the baron. And, my dear
Fatello, said he, detaining the banker, who, after shaking hands
with him, was about to leave the room, it is perhaps not necessary
to refer—at least not weigh upon—our conversation at last night's
masquerade. It might vex Mademoiselle Gonfalon—to learn that she
had been overheard—or—she might doubt your having heard, and
think I had been confiding to you a presumptuous and unfounded
belief of her partiality for myself. Women, you know, are susceptible
on these points; it might indispose her towards me, and lessen my
chance. In short, he added, with a smile, if you will be guided by
an ex-roué, now reformed, but who has some little experience of the
female heart, you will confine yourself to the communication of my
proposals, without reference to any thing past, and apply all your
eloquence to induce Mademoiselle Sebastiana to receive them as
favourably as yourself.
Fatello nodded knowingly.
75.
Ay, ay, saidhe, I see I need not despair of my ears. They do not
serve me so badly. But never fear, baron—I will know nothing,
except that you are desperately in love, and that your life depends
on your suit's success. That is the established formula, is it not?
When the baron—after escorting Fatello, in spite of his resistance, to
the door of the pavilion, where the banker's carriage awaited him—
re-entered the breakfast-room, the joyous and hopeful expression
his countenance had worn during the latter part of his conversation
with his visitor was exchanged for one of anxiety and doubt. Instead
of returning to the breakfast, of which he had scarcely eaten a
mouthful, he drew his arm-chair to the fire, threw himself into it,
and fell into a brown study. The attentive valet, who came in full of
concern for his master's interrupted meal, was sharply dismissed,
with an order to admit no callers. After a short time, however,
Steinfeld's cogitations apparently assumed a rosier hue. The wrinkles
on his brow relaxed their rigidity, he ceased to gnaw his mustache,
and at length a smile dawned upon his features, and grew till it
burst into a laugh. Something or other inordinately tickled the
baron's fancy; for he lay back in his chair and laughed heartily, but
silently, with the eyes rather than the mouth, for nearly a minute.
Then getting up, and lounging pensively through the room, he
indulged in a soliloquy of muttered and broken sentences, which,
like the secret cipher of a band of conspirators, were unintelligible
without a key. Their obscurity was increased by a style of metaphor
borrowed from the card-table, and which a man of such correct taste
as Steinfeld would doubtless have scrupled to employ in
conversation with any one but himself.
What an odd caprice of fate! he said. A strange turn in the game,
indeed! The card I most feared turns up trumps! It rather deranges
my calculations; but perhaps it is as good a card as the other.
Decidedly as sure a one. What certainty that yonder pedantic booby
is right in his prognostics? And then there was no avoiding it.
Provided, only, Fatello is silent about last night. If not, all is spoilt.
And if she makes a scene! Your Spanish dames are reputed fiery as
76.
Arabs; but Itake her for one of the milder sort—rather a pining than
a storming beauty. What if I were to miss both, by some infernal
quiproquo or other. Query, too, whether Sebastiana accepts; but I
think, with Fatello to back me, I need not fear much on that score. I
detect his motives. To your rich upstart, money is dirt compared with
descent, connexion, title. He would like to be an ambassador's
brother-in-law, the near connexion of a family dating from
Charlemagne—he, the man of nothing, with plebeian written on his
front. Upwards of half a million. Seven hundred thousand, I daresay.
I had reckoned on nearly double, and now I may lose both. Well, à
la grâce du diable. I will go take a gallop.
And in another half hour the aspirant to the hand and fortune of
Sebastiana Gonfalon was cantering round the Bois de Boulogne,
followed at the prescribed distance by Celestin, who, mounted on a
fine English horse, near sixteen hands high, bore no slight
resemblance to an ape exalted on an elephant.
CHAP. IV.—THE CAPTAIN'S ROOM.
The hotel of the Northern Eagle, situated in one of the most
respectable of the numerous small streets between the Rue St
Honoré and the Rue Neuve des Petits Champs, is one of several
hundred establishments of the class, scattered over Paris, and
which, although bearing the ambitious title of hotel, differ in no
essential respect from what in London are styled third or fourth-rate
lodging-houses. It is a tall, narrow, melancholy-looking edifice,
entered through an archway, which devours a great part of the
ground-floor, and is closed at night by a heavy coach-door, and in
the daytime by a four-foot palisade, painted a bright green, with a
gate in the middle, and a noisy bell that rings whenever the gate is
opened. Under the archway, and in the little paved court that
terminates it, there is always a strong smell of blacking in the
morning, and an equally strong smell of soup in the afternoon; the
former arising from the labours of Jean, a strapping, broad-
shouldered native of Picardy, who makes beds, cleans boots, and
carries water for the entire hotel; the latter emanating from a small,
77.
smoky den, notunlike a ship's caboose, where a dingy cookmaid
prepares the diurnal pot-au-feu for the mistress of the hotel, her son
and husband, and for a couple of pensioners, who, in consideration
of the moderate monthly payment of fifty francs each, are admitted
to share the frugal ragouts of Madame Duchambre's dinner-table. By
an architectural arrangement, common enough in old Paris houses,
and which seems designed to secure a comfortable gush of cold air
through the crevices of every door in the building, the foot of the
staircase is in the court, open to all weathers—a circumstance most
painful to Jean, who takes pride in the polish of his stairs, and is to
be seen, whenever his other avocations leave him a moment's
leisure, busily repairing, with a brush buckled on his foot, and a bit
of wax in a cleft stick, the damage done to their lustre by the muddy
boots of the lodgers. The hotel contains about five-and-twenty
rooms, all let singly, with the exception of the first floor, divided into
two appartemens of two rooms and a cupboard each, for which
Madame Duchambre obtains the extravagant rent of ninety and one
hundred francs per month. Above the first floor the rooms are of
various quality—from the commodious chamber which, by the
French system of an alcove for the bed, is converted in the daytime
into a very tolerable imitation of a parlour—to the comfortless attic,
an oven in summer, an ice-house in winter, dearly paid at five francs
a-week by some struggling artisan who works hard enough in the
day to sleep anywhere at night.
At the period referred to by this narrative, a room upon the third
floor of the hotel of the Northern Eagle was occupied, as might be
ascertained by inspection of a lithographed visiting card, stuck upon
the door with a wafer, by Godibert Carcassonne, captain in the 1st
African Chasseurs, known emphatically amongst the permanent
tenants of the hotel as The Captain. Not that military occupants
were a rarity under the wings of the Northern Eagle; captains were
common enough there—majors not very scarce—and it was upon
record that more than one colonel had occupied the yellow salon
upon the first floor. But none of these warriors bore comparison with
Captain Carcassonne in the estimation of Madame Duchambre, an
78.
elderly lady witha game leg, and a singularly plain countenance,
who had seen better days, and had a strong sense of the proprieties
of life. In general she professed no great affection for men of the
sword, whom she considered too much addicted to strong drink and
profane oaths, and who did not always, she said, respect la pudeur
de la maison. The captain, however, had completely won her heart—
not by any particular meekness or abstinence, for he consumed far
more cognac than spring water, had a voice like a deep-mouthed
mastiff, and swore, when incensed, till the very rafters trembled.
Nevertheless he had somehow or other gained her affections; partly,
perhaps, by the regularity with which, upon all his visits to Paris
during the previous fifteen years, he had lodged in her house and
paid his bills; partly, doubtless, by the engaging familiarity with
which he helped himself from her snuff-box, and addressed her as
Maman Duchambre.
It was eight o'clock at night, and, contrary to his wont, Captain
Carcassonne, instead of contesting a pool at billiards in his
accustomed café, or occupying a stall at his favourite Palais Royal
theatre, was seated in his room, alone, a coffee-cup and a bottle on
the table beside him, the amber mouthpiece of a huge meerschaum
pipe disappearing under his heavy dark mustache, smoking steadily,
and reading the Sentinelle de l'Armée. He was a powerful active
man, about forty years of age, with a red-brown complexion, martial
features, and a cavalier air, in whom Algerine climate and fatigues
had mitigated, if it had not wholly checked, that tendency to
corpulence early observable in many French cavalry officers, for the
most part a sedentary and full-feeding race. Of a most gregarious
disposition, no slight cause would have induced the captain to pass
in slow solitude those evening hours, which, according to his creed,
ought invariably, in Paris, to dance merrily by in the broad light of
gas, and in the excitement of a theatre or coffee-house. Neither was
it, in his eyes, a trifle that had placed him, as he expressed it, under
close arrest for the evening. He was paying a small instalment of a
debt of gratitude, which many would have held expunged by lapse
of time, but which Carcassonne still remembered and willingly
79.
acknowledged. Many yearspreviously—within a twelvemonth after
his promotion from a sergeantcy in a crack hussar regiment to a
cornetcy in a corps of chasseurs, newly formed for African service,
and in which he had since sabred his way to the command of a
troop—Godibert Carcassonne, when on leave of absence at Paris,
had been led, by thoughtlessness and by evil associates, rather than
by innate vice, into a scrape which threatened to blast his prospects
in the army, and consequently in life, and of his extrication from
which there was no possibility, unless he could immediately procure
five thousand francs. The sum was trifling, but to him it seemed
immense, for he estimated it by the difficulty of obtaining it. Driven
to desperation, thoughts of suicide beset him, when at that critical
moment a friend came to the rescue. By the merest chance, he
stumbled upon a former school-fellow, a native of the same
department as himself, and his accomplice in many a boyish frolic.
They had not seen each other for years. When Carcassonne was
taken by the conscription, his schoolmate had already departed to
seek fortune at Paris, the Eldorado of provincials, and there, whilst
the smart but penniless young soldier was slowly working his way to
a commission, he had taken root and prospered. He was not yet a
wealthy man, but neither was he a needy or niggardly one, for, on
hearing the tale of his friend's difficulties, he offered him, after a few
moments' internal calculation, the loan of the sum on which his fate
depended, and gruffly cut short the impetuous expression of
gratitude with which the generous offer was joyfully accepted. The
loan was in fact a gift, for when, some time afterwards, Carcassonne
remitted to his friend a small instalment of his debt, scraped
together by a pinching economy that did him honour, out of his
slender pay, the little draft was returned to him, with the words,
You shall pay me when you are colonel. And as all subsequent
attempts were met by the same answer, the money was still unpaid.
But never did loan bear better interest of gratitude. Carcassonne had
never forgotten the obligation, was never weary of seeking
opportunities of requiting it. These were hard to find, for his friend
was now a rich man, and there was little the dragoon could do for
him beyond choosing his horses, and giving his grooms valuable
80.
veterinary hints, derivedfrom his long experience of the chevaline
race in the stables of the 1st Chasseurs. Once only was he fortunate
enough to hear his benefactor slightingly spoken of at a public table
in Paris. That was a happy day for Carcassonne, and a sad one for
the offender, who was taken home a few hours afterwards with
pistol bullet in his shoulder.
The object of this devoted attachment on the part of the rough but
honest-hearted soldier, was not insensible to the sincerity and value
of such friendship, and returned it after his own fashion,—that is to
say, somewhat as the owner of a noble dog permits its
demonstrations of affection, and requites them by an occasional
caress. When Carcassonne came to Paris, which he did as often as
he could get leave of absence from his duties in Africa, his first visit
was always for his benefactor, who invariably got up a dinner for him
—not at his own house, which the dragoon would have considered a
tame proceeding, but at some renowned restaurant—a regular
bamboche, as the African styled it, where champagne corks flew and
punch flamed from six in the evening till any hour after midnight.
Then, the civilian's occupations being numerous, and his sphere of
life quite different from that of the soldier, the two saw but little of
each other, except through a casual meeting in the rich man's
stables, or on the boulevard, or when—but this was very rare—
Carcassonne was surprised in his room, at the Northern Eagle, by an
unexpected but most welcome visit from his friend, come to smoke a
passing cigar, and have ten minutes' chat over boyish days and
reminiscences.
These visits were a great treat to the captain; and it was the
anticipation of one of them that now kept him in his room. To his
astonishment, he had received that morning a note from his friend,
requesting him to remain at home in the evening, as he would call
upon and crave a service of him. Carcassonne was delighted at the
intimation, and not feeling quite certain when evening might be said
to begin, he shut himself up in his room at four o'clock, ordered in
dinner from a neighbouring traiteur, sipped his coffee in contented
81.
solitude, and nowawaited, with the dutiful patience of a soldier on
sentry, the promised coming of his friend. At last a cough and a
heavy footstep were heard upon the stairs; the captain took up a
candle, opened the door, and, stepping out into the gloomy corridor,
the light fell upon the tall ungainly figure, and sullen features, of
Sigismund Fatello.
Come in, my dear fellow, cried Carcassonne in his stentorian tones,
and with a soldier's oath. I've expected you these three hours.
What—wet? Snow? Come to the fire, and take a sup of cognac till
the punch is made.
It snowed heavily outside, and the banker's upper coat had caught a
few large flakes in crossing the court. He heeded them not, but
putting down, untasted, the glass of brandy handed to him by the
captain, he took a chair, and motioned Carcassonne to another.
What the deuce is the matter with you, Sigismund? said the
captain, looking hard at his friend. Are you ill?
Better than I have for a long time been. Fresh from a wedding.
Oho! said Carcassonne. I thought you had not put on full dress to
visit your old comrade in his den at the Northern Eagle. And whose
wedding was it?
A singular one, replied the banker, parrying the question.
Strangely brought about, certainly. Would you like to hear its
history, Carcassonne?
By all means, said the captain, who always liked whatever Fatello
proposed. But the business you came about?—you said I could do
something for you. What is it?
Plenty of time for that. It will keep. Let me tell you of this
marriage.
Delighted to listen, said Carcassonne, settling himself in his chair,
and filling his pipe from a huge embroidered bag, once the property
82.
of an ArabianEmir's lady, but which a razzia had degraded into a
receptacle for tobacco.
You must know, then, Carcassonne, said Fatello, that a friend of
mine, named Oliver, a man of middle age, more calculated to shine
in a counting-house than in a boudoir, was fool enough, not very
long ago, to fall in love with a beautiful girl, twenty years younger
than himself; and as he was rich, and her father avaricious, the
marriage was brought about, although not altogether with her good
will.
Bad, quoth the captain, between two puffs of his pipe. An
unwilling bride is apt to prove a sour wife.
Once married, continued Fatello, without heeding his friend's
interruption, Oliver, who knew he had not his wife's love, spared no
pains to obtain her friendship. He was not such a man, either by
person, manners, or temper, as women are apt to fancy; but, to
atone for his deficiencies, he covered her with gold, was the slave of
her caprices, forestalled her slightest wish. Her amusement and
happiness were the whole study of his life; and after a while his
efforts seemed crowned with success. She treated him as a friend,
and appeared contented with her lot. This was all he had dared to
hope, and, having attained this, he was happy. His existence, from
boyhood upwards, had been agitated and laborious, but riches had
rewarded his toils, and he could now look forward to a long period of
happiness and repose. At the very moment he indulged these visions
of a bright future, a single word, whispered in his ear by a physician
of high repute, crumbled the entire fabric. That word was
Consumption, and when he heard it he knew his doom was sealed.
His father, his elder brother, his sisters, all had been carried off, in
the prime of their strength, by the insidious disease, whose germ,
implanted in their system before they saw the light, was ineradicable
by the resources of art. The shock was severe—it could not be
otherwise—for most of the things were his for which men prize life.
But he was no poltroon, to pine at the approach of death; and he
nerved himself to meet like a man his inevitable fate. Although with
83.
scarce a shadowof hope, he neglected no means of combating the
deadly malady; and, enjoining secrecy to his physician, he concealed
from every one his belief that his days were numbered and his race
wellnigh run. He was calm and resigned, if not hopeful, when he one
day received a letter that chilled his very soul. His wife, it told him,
loved another, whom she would meet that night at a masquerade.
Although anonymous, its indications were so precise, that Oliver,
spurred by fiercest jealousy, disguised himself and went secretly to
the ball. There he discovered his wife, in the company of a foreign
fopling, who, for some time previously, had been a frequent visitor
at his house. He kept near them, occasionally catching a sentence
confirmatory of his suspicions, until they withdrew from the crowd,
and sought a retired nook, where to converse uninterrupted. He
found means to secrete himself in their vicinity, and overheard—no
evidence of his dishonour, for then he had stabbed them where they
stood—but words whence he gathered the existence of the most
heartless, perfidious, and cold-blooded calculation.
The wife of his bosom, to gain whose affection he had squandered
millions, and changed his very nature, impatiently awaited his death
to bestow her hand, and the fortune he should bequeath her, on the
smooth-tongued seducer whose arts had beguiled her. The secret of
his fatal malady had been divulged by the physician, to whom alone
it was known, in the hearing of this foreign adventurer, who, ever
upon the watch to redeem his broken fortunes by a wealthy
marriage, profited by the disclosure. He obtained an introduction to
Oliver's house, and applied every art and energy to gain his wife's
affections. He was but too successful. She listened to his
protestations, and on learning her husband's impending death,
pledged herself to become his, when she should be released by it
from ties she abhorred. All this, and more, Oliver gathered from their
conversation, to which he had the courage to listen to the end,
although each sentence went to his heart like a stab, leaving in the
wound the venom of hate and jealousy, to rankle there until the
latest moment of his life. What had you done, Carcassonne, had you
been in his place?
84.
Pardieu! said thecaptain, who had listened with profound
attention, and great expenditure of smoke, to his friend's narrative;
I can hardly say, Sigismund. If I had kept my hands off the butterfly
scoundrel when I heard him courting my wife, I should have
followed him when he had had his chat out, and requested the
pleasure of crossing swords with him at his earliest convenience;
and had I got one good cut at him, he should not have needed
another. What did your friend?
Very nearly what you have said. He went home and destroyed his
will, and made another. Then he sought his enemy, to challenge him
to an instant encounter. The mean villain denied his treachery, and
swore that her to whom his vows of love were addressed was not
Oliver's wife, but his sister-in-law. Oliver well knew this to be a lie,
but he affected to believe he had been deceived by similarity of
dress and imperfect hearing, for the subterfuge had suddenly
suggested to him a sure means of punishing his faithless wife, and
defeating her seducer's aim. He declared himself willing to aid the
views of the foreigner—one Baron Steinfeld, an Austrian of high
family, but ruined fortunes—and to urge his sister-in-law to accept
his hand. Disagreeably surprised at such willingness, where he had
wished and expected opposition, Steinfeld strove to recede, but
found extrication impossible from the trap he had rushed into. Finally
he was compelled to yield; the less unwillingly because the bride
thus given him was not without fortune, which Oliver exaggerated,
the better to allure him. So that, when Oliver left him, it was to
convey his formal proposals to the lady, who was nothing loath, and
to-day they were married.
To-day! exclaimed Carcassonne. This, then, is the wedding you
come from. And what said Madame Oliver?
What could she say? Made all the secret opposition she could, no
doubt; and then, finding it in vain, for her sister seemed as much
fascinated by the Austrian Lothario as she was herself, she took ill
and kept her bed. It needed all her woman's pride, and her fear of
85.
Welcome to ourwebsite – the perfect destination for book lovers and
knowledge seekers. We believe that every book holds a new world,
offering opportunities for learning, discovery, and personal growth.
That’s why we are dedicated to bringing you a diverse collection of
books, ranging from classic literature and specialized publications to
self-development guides and children's books.
More than just a book-buying platform, we strive to be a bridge
connecting you with timeless cultural and intellectual values. With an
elegant, user-friendly interface and a smart search system, you can
quickly find the books that best suit your interests. Additionally,
our special promotions and home delivery services help you save time
and fully enjoy the joy of reading.
Join us on a journey of knowledge exploration, passion nurturing, and
personal growth every day!
ebookbell.com










![vi Preface
biologists. We highly recommend the reader study this material in more detail by
consulting one of the many excellent books devoted primarily to neuroscience. Such
books include those by Kandel et al. [144] and Johnston and Wu [139].
We have also tried to provide background material on dynamical systems theory,
including phase plane methods, oscillations, singular perturbations, and bifurcation
analysis. An excellent way to learn this material is by using it, together with com-
puter simulations, to analyze interesting, concrete examples. The only prerequisites
are a basic knowledge of calculus, knowledge of a little linear algebra (matrices,
eigenvalues), and understanding of some basic theory of ordinary differential equa-
tions. Much of the mathematics is at the level of Strogatz [255].
The book is organized from the bottom up. The first part of the book is con-
cerned with properties of a single neuron. We start with the biophysics of the cell
membrane, add active ion channels, introduce cable theory, and then derive the
Hodgkin–Huxley model. Chapter 2 is concerned with the basic properties of den-
drites. We then introduce dynamical systems theory, using a simple neuron model to
illustrate the basic concepts. We return to the biology in Chap. 4, where we discuss
the variety of ion channels which have been found in neurons. Chapters 5 and 6
are devoted to bursting oscillations and propagating action potentials, respectively.
Here, we use many of the dynamical systems techniques to describe mechanisms
underlying these behaviors. The second part of the book is concerned with neu-
ronal networks. In Chap. 7, we describe synaptic channels, which are the primary
way that neurons communicate with each other. Chapters 8 and 9 discuss two dif-
ferent approaches for studying networks. First, we assume weak coupling and use
phase-response methods. We then demonstrate how one can analyze firing patterns
in neuronal networks using fast/slow analysis. In Chap. 10, we discuss the role of
noise in neuron models. Here, we briefly introduce the reader to the mathematical
theory of stochastic differential equations. Finally, in Chaps. 11 and 12 we discuss
firing rate models and spatially distributed networks.
There is far more material in this book than could be covered in a one-semester
course. Furthermore, some of the material is quite advanced. A course in computa-
tional neuroscience slanted toward mechanisms and dynamics could easily be made
out of the first five chapters along with Chap. 7. These chapters would cover most
of the basics of single-cell modeling as well as introduce students to dynamical sys-
tems. The remainder of such a course could include selections from Chaps. 8–12.
For example, Chap. 11 contains firing rate models, with many applications provided
in Sect. 11.3. Parts of Chap. 12 could comprise the remainder of the course.
For more mathematically inclined students, the elementary dynamics chapter
(Chap.3) could be skipped and the more technical chapters could be emphasized.
There is lovely nonlinear dynamics in Chaps. 5, 6, 8, and 9, which along with the
earlier chapters could form the core of a mathematical neuroscience course.
There are several recent books that cover some of the same material as in
the present volume. Theoretical Neuroscience by Dayan and Abbott [53] has a
broader range of topics than our book; however, it does not go very deeply into
the mathematical analysis of neurons and networks, nor does it emphasize the
dynamical systems approach. A much more similar book is Dynamical Systems](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-11-2048.jpg)
![Preface vii
in Neuroscience by Izhikevich [136]. This book emphasizes the same approach
as we take here; however, the main emphasis of Dynamical Systems in Neu-
roscience is on single-neuron behavior. We cover a good deal of single-neuron
biophysics, but include a much larger proportion of theory on systems neuro-
science and applications to networks. There are many specific models and equa-
tions in this text. The forms of these models and their parameters are available at
http://www.math.pitt.edu/bard/bardware/neurobook/allodes.html.
Pittsburgh, PA G. Bard Ermentrout
Columbus, OH David H. Terman](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-12-2048.jpg)















![1.3 The Goldman–Hodgkin–Katz Equation 7
Given several ionic species, the total current is just a sum of the individual cur-
rents. This is a consequence of assumption 3, which says that the ions do not interact.
Suppose there are three permeable ions, KC
, NaC
, and Cl
with corresponding
currents, IK; INa, and ICl: At equilibrium, the total current, I D IK C INa C ICl,
vanishes; that is, I D 0: The potential at which this occurs is
VM D
RT
F
ln
PKŒKC
out C PNaŒNaC
out C PClŒCl
in
PKŒKCin C PNaŒNaCin C PClŒClout
; (1.4)
where the Pj ’s are the permeabilities of each of the three ionic species. This is a
generalization of the Nernst equilibrium discussed above and is called the Goldman–
Hodgkin–Katz (GHK) equation. With one species, the equation reduces to the
Nernst potential. For example, in the squid axon, the ratios of the permeabilities,
at rest, are PK W PNa W PCl D 1 W 0:03 W 0:1. The ion concentrations inside the cell
are, respectively, for KC
, NaC
, and Cl
, 400, 50, and 40 mM, whereas outside the
cell they are 10, 460, and 540 mM. Thus, at room temperature, the equilibrium or
resting potential is 74 mV.
Table 1.1 Typical ion concentrations in cells (from Johnston and Wu [139])
Equilibrium potential (mV),
Ion Inside (mM) Outside (mM) Ei D RT
zF
ln
ŒCout
ŒCin
Frog muscle T D 20ı
C
KC
124 2.25 58 log 2:25
124
D 101
NaC
10.4 109 58 log 109
10:4
D C59
Cl
1.5 77.5 58 log 77:5
1:5
D 99
Ca2C
104
2.1 29 log 2:1
104 D C125
Squid axon T D 20ı
C
KC
400 20 58 log 20
400
D 75
NaC
50 440 58 log 440
50
D C55
Cl
40–150 560 58 log 560
40150
D 66 to 33
Ca2C
104
10 29 log 10
104 D C145
Mammalian cell T D 37ı
C
KC
140 5 62 log 5
140
D 89:7
NaC
5–15 145 62 log 145
515
D C90 .C61/
Cl
4 110 62 log 110
4
D 89
Ca2C
104
2.5–5 31 log 2:55
104 D C 136 .C145/](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-28-2048.jpg)











![1.8 Voltage-Gated Channels 19
gate is open or closed depends on the membrane potential. The gate model can be
summarized by the diagram
C
˛.V /
•
ˇ.V /
O; (1.33)
where C and O correspond to the closed and open states, respectively, and ˛.V /
and ˇ.V / are the voltage-dependent rate constants at which a gate goes from the
closed to the open and from the open to the closed states, respectively. If we let m
be the fraction of open gates, then 1 m is the fraction of closed gates, and, from
the law of mass action,
dm
dt
D ˛.V /.1 m/ ˇ.V /m D .m1.V / m/= .V /; (1.34)
where
m1.V / D
˛.V /
˛.V / C ˇ.V /
and .V / D
1
˛.V / C ˇ.V /
: (1.35)
It is easy to solve this equation if V is constant. The solution starting at m.0/ is
m.t/ D m1.V / C .m.0/ m1.V //et=.V /
:
Note that the solution approaches the steady-state m1.V / at a rate determined by
the time constant .V /.
One must obtain expressions for the voltage-dependent rate constants ˛ and ˇ.
In the Hodgkin–Huxley model, these functions were derived by fitting the data.
Borg-Graham [17] and others have suggested a simple formulation based on thermo-
dynamics. The idea is that the probability of opening or closing a channel depends
exponentially on the potential. Thus,
˛.V / D A˛ exp.B˛V / and ˇ.V / D Aˇ exp.Bˇ V /: (1.36)
From this, we find that
m1.V / D
1
1 C exp..V Vh/=Vs/
;
where Vh and Vs are constants. We leave as an exercise the calculation of these
constants in terms of the constants A and B. The time constant, .V /, will generally
be a skewed bell-shaped function of V: If Bˇ D B˛, then .V / is a hyperbolic
secant.](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-40-2048.jpg)
![20 1 The Hodgkin–Huxley Equations
1.9 Hodgkin–Huxley Model
We are now ready to derive the Hodgkin–Huxley model for the propagation of an
action potential along the squid’s giant axon. We view the axon as a cylinder of fixed
radius, a, so the membrane potential depends on the spatial variable x and time t.
Here, we assume there are voltage-gated KC
and NaC
channels and a leak current.
Then balancing currents, as in (1.23), we have
IL D Icap C Iion (1.37)
or, using (1.6) and (1.24),
a
2rL
@2
VM
@x2
D cM
@VM
@t
C IK C INa C IL: (1.38)
If each ionic current is ohmic, then this can be written as
cM
@VM
@t
D
a
2rL
@2
VM
@x2
gK.VM EK/ gNa.VM ENa/ gL.VM EL/: (1.39)
To complete the model, we need to describe how one computes the membrane con-
ductances gK; gNa, and gL. Note that the voltage-gated conductances gK and gNa
change with time during an action potential.
Hodgkin and Huxley used two experimental methods to separate the ionic cur-
rents and compute how the KC
and NaC
conductances depend on voltage. The first
was a simple feedback circuit called the voltage clamp that allows the experimenter
to hold the membrane potential at a constant or holding level VC. The voltage clamp
does so by injecting a current into the axon that is equal and opposite to the cur-
rent flowing through the voltage-gated channels. Electrical details can be found in
the book by Johnston and Wu [139]. Note that the voltage clamp separates the total
membrane current into its ionic and capacitive components. Recall that the capaci-
tive current satisfies Icap D CMdVM=dt. If the membrane potential is fixed at some
constant, then the capacitive current must be zero. Moreover, the total current can be
made spatially uniform by inserting a highly conductive axial wire inside the fiber;
the axon is then said to be space-clamped. In this case, @2VM
@x2 D 0: It then follows
that any changes in current must be due to either the leak or the opening and closing
of voltage-gated membrane channels.
We first consider how the voltage clamp can be used to determine the leak
conductance, gL. Note that most of the voltage-gated channels are closed at rest.
Moreover, if we hyperpolarize the cell, then we may assume all of the voltage-gated
channels are closed. It follows that if the membrane potential is clamped at some
sufficiently strong hyperpolarized level, then the total current is given by the leak;
that is,
IM gL.VC EL/:
From this equation, we can easily solve for gL.](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-41-2048.jpg)

![22 1 The Hodgkin–Huxley Equations
−60
−40
−20
0
20
40
V
C
(mV)
0
10
20
30
0 1 2 3 4 5 6 7
0
20
40
time (msec)
gK
gNa
Fig. 1.9 Numerically computed voltage-clamp experiment. The membrane potential is stepped to
different values and the resulting KC
and NaC
conductances are computed
begin to close before the depolarization is turned off, whereas the KC
channels
remain open as long as the membrane is depolarized. This suggests the NaC
channel
can exist in three states: resting, activated, and inactivated. When the cell is depo-
larized, the NaC
channels switch from the resting (closed) to the activated (open)
state. If the depolarization is maintained, then the channel switches to the inactivated
(closed) state.
A physical interpretation of the NaC
channel is shown in Fig. 1.10. There are
two gates in the NaC
channel: a fast one (the activation gate), represented by the
line, and a slow one (the inactivation gate), represented by the ball. Both gates must
be open for the channel to conduct NaC
ions. At rest, the activation gate is closed
and the inactivation gate is open. When the membrane is depolarized, the activation
gate opens, which allows NaC
into the cell. The inactivation gate (ball) closes at
the higher potential, so the flow of NaC
is transient. Hodgkin and Huxley used a
more complicated voltage-clamp protocol, first stepping to a fixed voltage and then
applying brief voltage steps to probe the fast activation and slow inactivation gates.
Details can be found in [144].
Using the voltage-clamp data, Hodgkin and Huxley derived expressions for the
KC
and NaC
conductances. They proposed that
gK D N
gKn4
and gNa D N
gNam3
h; (1.41)
where N
gK and N
gNa are maximum conductances and n, m, and h are gating variables
that take values between 0 and 1. Hence, n4
represents the probability that a KC](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-43-2048.jpg)




![1.11 Bibliography 27
V increases
Na+ channels
activate: m
Na+ enters
cell
Na+ channels
inactivate: h
K+ channels
activate: n
Na+ deactivates: m
Na+ deinactivates: h
K+ deactivates: n
EK
ENa
Vrest
V
time
Fig. 1.15 Mechanisms underlying the action potential
It follows that eventually, the KC
current dominates and the membrane potential
must fall back toward the KC
Nernst potential. This corresponds to the downstroke
of the action potential.
After the action potential (Fig. 1.15), the cell is hyperpolarized with m1 0;
n1 0, and h1 1. After some time, m, n, and h approach their steady-state
values and the cell returns to rest.
1.11 Bibliography
There are many standard neuroscience textbooks that present the biological as-
pects covered in this chapter in much more detail. These textbooks include those
by Kandel et al. [144], Hille [122], and Martin [192]. The reader is also highly
recommended to look at Hodgkin and Huxley’s original papers [124]. A review
of these papers, along with a short review of the history leading up to them, is
given in Rinzel [229]. Excellent textbooks which emphasize modeling and quanti-
tative approaches are those of Johnston and Wu [139], Koch [156], Jack et al. [137],
Izhikevich [136], and Dayan and Abbott [54]. Keener and Sneyd [148] and Fall
et al. [83] give detailed introductions to mathematical aspects of cellular biophysics.](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-48-2048.jpg)
![28 1 The Hodgkin–Huxley Equations
1.12 Exercises
1. Suppose the external potassium in a mammalian cell is increased by a factor of
10. What is the new value of EK?
2. At 10ı
C a cell contains 80 mM sodium inside and has only 100 mM sodium
outside. What is the equilibrium potential for sodium?
3. Compute the resting potential for the mammalian cell using the same perme-
abilities as were used for the squid axon and the ion concentrations listed in
Table 1.1.
4. Derive the Nernst equation (1.2) from the Nernst–Planck equation by setting the
current to zero and integrating with respect to x across the membrane.
5. Compute the calcium equilibrium potential for a mammalian cell assuming that
the extracellular concentration is 5 mM and the intracellular concentration is
104
mM.
6. Complete the derivation the constant-field equation (1.3) from the linear Nernst–
Planck equation.
7. Derive the GHK equation (1.4) from the constant-field equation.
8. Consider the GHK equation and plot the I–V relation for different values of
the inside and outside concentrations. Show that for ŒCout ŒCin (respectively
ŒCout ŒCin) the I–V plot is concave down (respectively up).
9. Consider a passive, spherical cell with radius 0:003 cm2
, a resting membrane
potential of 65 mV, a membrane capacitance of 1F=cm2
, and a membrane
resistance of RM D 700 cm2
. Suppose the cell is injected with an applied
current of 5 nA/m2
for 2 s and then the current is turned off. What is the mem-
brane potential at t D 1; t D 2, and t D 3?
10. Suppose a passive axon has a diameter of 0.5 mm, a resting membrane resis-
tance of RM D 700 cm2
, and a transmembrane resistance of RL D 30 cm.
Compute the space constant. If the axon is 5 cm long and one end of the axon is
held at 120 mV above rest, then what is the potential at the other end?
11. (Johnston and Wu [139], page 12) The membrane capacitance of a typical cell
is 1 F=cm2
and the concentration of ions inside and outside the cell is about
0.5 M. Calculate the fraction of uncompensated ions on each side of the mem-
brane required to produce 100 mV in a spherical cell with a radius of 25 m.
12. Numerically solve the Hodgkin–Huxley equations. Start the system at rest and,
at some later time, inject an applied current to generate an action potential. Plot
the time courses of the NaC
and KC
conductances, as well as the gating vari-
ables m, h and n.
13. Numerically perform space-clamp experiments. That is, start the Hodgkin–
Huxley model at rest and, at some later time, change the membrane potential
and keep it as some “clamped” level. Plot the NaC
and KC
conductances for
when the membrane potential is stepped to different values.](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-49-2048.jpg)



![32 2 Dendrites
and thus the ratio of the coupled to the uncoupled input resistance is
Rinput;coupled
Rinput;uncoupled
D 1
rM
r C 2rM
I
that is, the input resistance decreases. To get the same increment in potential, the
current required for the coupled system is more than that required for the uncoupled
system because some current is drained away by the second compartment.
In a similar way, we can derive a compartmental model for a general treelike
structure. A general algorithm for computing the correct equations is
For each cylinder, j, with radius and length aj and Lj in micrometers,
compute the surface area, Aj D 2aj Lj , and the axial resistance factor,
Qj D Lj =.a2
j /:
The membrane capacitance is Cj D cj Aj 108
and the membrane resistance
is Rj D .rMj =Aj / 108
.
The coupling resistance between compartments j and k is Rjk D rL
2
.Qj C
Qk/ 104
.
The equations are then
Cj
dVj
dt
D
Vj
Rj
C
X
k connected j
Vk Vj
Rjk
C Ij :
The factors of 10˙8
and 104
are the conversion from micrometers to centimeters.
For example, consider a two-compartment model with (1) compartment 1 having a
length of 200 m and radius of 30 m and (2) compartment 2 having a length of
20 m and radius of 20 m. Then, R1 D 2:65 107
; C1 D 3:77 1010
F,
R2 D 3:98 108
; C2 D 2:52 1011
F, and Rlong D 4:34 104
: Thus,
10
dV1
dt
D V1 C 611.V2 V1/; 10
dV2
dt
D V2 C 9;181.V2 V1/;
where time is in milliseconds and the coupling coefficients are dimensionless. Note
how the ratio of the coupling strengths is the same as the reciprocal of the area ratios.
The bigger compartment has a much greater effect on the smaller compartment than
vice versa.
We also remark that the standard units used in most compartmental models are
microfarads per square centimeter, millisiemens per square centimeter, and mi-
croamperes per square centimeter for the capacitance, conductance, and applied
current, respectively. Experimentalists do not generally know the current density,
but only the total current injected. Typical currents injected into a cell are of the
order of less than 1 nA.
To generate arbitrary compartmental models, one needs only to compute the
length, diameter, and the connectivity of the cylinders that make up the dendritic
tree. The program NEURON [33] enables an experimentalist to input a digitized
picture of a neuron and the program will automatically produce a compartmental
model of the neuron by linking together many cylinders.](https://image.slidesharecdn.com/1268387-250525203558-c146807b/75/Mathematical-Foundations-Of-Neuroscience-1st-Edition-G-Bard-Ermentrout-53-2048.jpg)