Base Editors Methods And Protocols 1st Edition Sangsu Bae Beomjong Song Editors
Base Editors Methods And Protocols 1st Edition Sangsu Bae Beomjong Song Editors
Base Editors Methods And Protocols 1st Edition Sangsu Bae Beomjong Song Editors
Base Editors Methods And Protocols 1st Edition Sangsu Bae Beomjong Song Editors
Base Editors Methods And Protocols 1st Edition Sangsu Bae Beomjong Song Editors
1.
Base Editors MethodsAnd Protocols 1st Edition
Sangsu Bae Beomjong Song Editors download
https://ebookbell.com/product/base-editors-methods-and-
protocols-1st-edition-sangsu-bae-beomjong-song-editors-48808756
Explore and download more ebooks at ebookbell.com
2.
Here are somerecommended products that we believe you will be
interested in. You can click the link to download.
Base Excision Repair Pathway Methods And Protocols Methods In
Molecular Biology 2701 1st Ed 2023 Kishor K Bhakat Editor
https://ebookbell.com/product/base-excision-repair-pathway-methods-
and-protocols-methods-in-molecular-biology-2701-1st-ed-2023-kishor-k-
bhakat-editor-51569302
Fragmentbased Approaches In Drug Discovery Methods And Principles In
Medicinal Chemistry 1st Edition Wolfgang Jahnke
https://ebookbell.com/product/fragmentbased-approaches-in-drug-
discovery-methods-and-principles-in-medicinal-chemistry-1st-edition-
wolfgang-jahnke-2164782
Methods And Algorithms For Molecular Dockingbased Drug Design And
Discovery Siavoush Dastmalchi
https://ebookbell.com/product/methods-and-algorithms-for-molecular-
dockingbased-drug-design-and-discovery-siavoush-dastmalchi-47711020
Level Set And Pde Based Reconstruction Methods In Imaging Cetraro
Italy 2008 Editors Martin Burger Stanley Osher 1st Edition Martin
Burger
https://ebookbell.com/product/level-set-and-pde-based-reconstruction-
methods-in-imaging-cetraro-italy-2008-editors-martin-burger-stanley-
osher-1st-edition-martin-burger-4411944
3.
Physiologicallybased Pharmacokinetic ModelingMethods And Applications
In Toxicology And Risk Assessment 1st Edition Jeffrey Fisher Editor
https://ebookbell.com/product/physiologicallybased-pharmacokinetic-
modeling-methods-and-applications-in-toxicology-and-risk-
assessment-1st-edition-jeffrey-fisher-editor-11171434
Exploring Digital Technologies For Artbased Special Education Models
And Methods For The Inclusive K12 Classroom 1st Edition Rick L Garner
Editor
https://ebookbell.com/product/exploring-digital-technologies-for-
artbased-special-education-models-and-methods-for-the-
inclusive-k12-classroom-1st-edition-rick-l-garner-editor-34998152
Riskbased Structural Evaluation Methods Best Practices And Development
Of Standards Ming Liu Editor Bruce R Ellingwood Editor Michel Ghosn
Editor
https://ebookbell.com/product/riskbased-structural-evaluation-methods-
best-practices-and-development-of-standards-ming-liu-editor-bruce-r-
ellingwood-editor-michel-ghosn-editor-23990428
Cases On Digital Gamebased Learning Methods Models And Strategies 1st
Edition Nicola Whitton Editor
https://ebookbell.com/product/cases-on-digital-gamebased-learning-
methods-models-and-strategies-1st-edition-nicola-whitton-
editor-37757916
Evidencebased Policy Making In The Social Sciences Methods That Matter
Gerry Stoker Editor Mark Evans Editor
https://ebookbell.com/product/evidencebased-policy-making-in-the-
social-sciences-methods-that-matter-gerry-stoker-editor-mark-evans-
editor-51808890
M E TH O D S I N M O L E C U L A R B I O L O G Y
Series Editor
John M. Walker
School of Life and Medical Sciences
University of Hertfordshire
Hatfield, Hertfordshire, UK
For further volumes:
http://www.springer.com/series/7651
7.
For over 35years, biological scientists have come to rely on the research protocols and
methodologies in the critically acclaimed Methods in Molecular Biology series. The series was
the first to introduce the step-by-step protocols approach that has become the standard in all
biomedical protocol publishing. Each protocol is provided in readily-reproducible step-by-
step fashion, opening with an introductory overview, a list of the materials and reagents
needed to complete the experiment, and followed by a detailed procedure that is supported
with a helpful notes section offering tips and tricks of the trade as well as troubleshooting
advice. These hallmark features were introduced by series editor Dr. John Walker and
constitute the key ingredient in each and every volume of the Methods in Molecular Biology
series. Tested and trusted, comprehensive and reliable, all protocols from the series are
indexed in PubMed.
8.
Base Editors
Methods andProtocols
Edited by
Sangsu Bae and Beomjong Song
Seoul National University College of Medicine, Seoul, Republic of Korea
Preface
The emergence ofthe CRISPR-Cas system has revolutionized many research areas, includ-
ing life sciences, biotechnology, and medicine, due to its ease of use and precise editing
ability. However, several studies have reported on safety concerns associated with the use of
CRISPR nucleases, such as the p53-mediated DNA response, programmed cell death, large
deletions, and/or chromosomal rearrangements that occur during the process of DNA
repair in cells. To bypass this issue, one breakthrough was achieved with the development
of base editing technology. DNA base editors (BEs), including cytosine base editors (CBEs)
and adenine base editors (ABEs), can convert target nucleotides with a single-nucleotide
resolution without generating DNA double-strand breaks (DSBs) or requiring donor DNA
templates. Due to such advantages, diverse variants of BEs are being widely used.
The aim of this Methods in Molecular Biology (MIMB) book is to introduce BEs and their
versatile applications to provide readers with a general understanding of BEs. Thanks to
contributions by excellent scientists, the book was written in four parts. The first part
(Chapters 1, 2, 3, and 4) introduces the history and diverse variants of BEs, and useful
tools for target selection, prediction of base editing efficiencies, and analysis of editing
outcomes/genome-wide off-target sites. The second part (Chapters 5, 6, 7, 8, and 9)
describes experimental applications of BEs for disease modeling in mammalian cells and
generating mutagenic mice. The third part (Chapters 10, 11, 12, 13, and 14) describes
in vivo/ex vivo therapeutic base editing strategies, which cover delivery methods of
BE-encoded DNA plasmids, mRNAs, or ribonucleoproteins through viruses or nonviral
lipid nanoparticles. The last part (Chapters 15, 16, and 17) describes the use of BEs in plants
and bacteria.
Each chapter describes the experimental process step-by-step with a list of all materials
required for the experiments so that readers can follow it easily even though they do not
have any prior experience with the experiments. In addition, the Note sections at the end of
the protocols provide precautions and small tips to minimize trial and error. In conclusion,
this collection of chapters will be a good guide for those who want to proceed with their
work using BEs in various fields.
We would like to thank all the authors, Dr. John M. Walker, the series editor, and
everyone involved in the publication of the book.
Seoul, Republic of Korea Sangsu Bae
Beomjong Song
v
11.
Contents
Preface . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . v
Contributors. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ix
PART I OVERVIEW OF BES, DIVERSE VARIANTS OF BES, AND COMPUTATIONAL
TOOLS FOR BES
1 Introduction and Perspectives of DNA Base Editors. . . . . . . . . . . . . . . . . . . . . . . . . 3
Beomjong Song and Sangsu Bae
2 Web-Based Computational Tools for Base Editors . . . . . . . . . . . . . . . . . . . . . . . . . . 13
Gue-Ho Hwang and Sangsu Bae
3 Prediction of Base Editing Efficiencies and Outcomes Using DeepABE
and DeepCBE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
Jinman Park and Hui Kwon Kim
4 Profiling Genome-Wide Specificity of dCpf1 Cytidine Base Editors
Using Digenome-Seq . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
Daesik Kim
PART II APPLICATION OF BES FOR DISEASE MODELING
5 Base Editing of Human Hematopoietic Stem Cells. . . . . . . . . . . . . . . . . . . . . . . . . . 43
Jing Zeng, Gabriele Casirati, My Anh Nguyen,
Pietro Genovese, and Daniel E. Bauer
6 A/C Simultaneous Conversion Using the Dual Base Editor
in Human Cells. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
Xiaohui Zhang, Yuting Guan, and Dali Li
7 Functional Analysis of Variants in BRCA1 Using CRISPR Base Editors. . . . . . . . 73
Ji-Eun See and Yongsub Kim
8 Use of the Representative Base Editing Tool Target-AID
to Introduce Pathogenic Mutations into Mice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87
Hiroki Sasaguri
9 Targeted Mutagenesis in Mice Using a Base Editor . . . . . . . . . . . . . . . . . . . . . . . . . 99
Tae Yeong Jeong, Soo-Yeon Lim, Je Kyung Seong,
and Kyoungmi Kim
PART III THERAPEUTIC BASE EDITING STRATEGIES
10 Heterologous Expression and Purification of a CRISPR-Cas9-Based
Adenine Base Editor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
Seu-Na Lee, Hong-Su Jang, and Jae-Sung Woo
vii
12.
viii Contents
11 DeliveringBase Editors In Vivo by Adeno-Associated
Virus Vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 135
Colin K. W. Lim, Angelo J. Miskalis, Pablo Perez-Pinera,
and Thomas Gaj
12 The Delivery of ABE mRNA to the Adult Murine Liver
by Lipid Nanoparticles (LNPs) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159
Dong Yang and Chun-Qing Song
13 Ex Vivo Base Editing Therapy with Chemically Derived
Hepatic Progenitors. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
Yohan Kim, Jaemin Jeong, and Dongho Choi
14 Application of Base Editor-Mediated Genome Editing
in Mouse Retina . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 179
Jun Wu, Chang Sik Cho, Dong Hyun Jo,
and Jeong Hun Kim
PART IV USE OF BES IN PLANTS AND BACTERIA
15 High-Throughput Base Editing-Mediated Artificial Evolution
Streamlines Trait Gene Identification in Rice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 191
Fang Yan, Man Yu, Meixia Wang, and Huanbin Zhou
16 Heritable Virus-Induced Genome Editing (VIGE)
in Nicotiana attenuata . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 203
Hyeonjin Kim, Youngbin Oh, Eunae Park, Moonyoung Kang,
Yuri Choi, and Sang-Gyu Kim
17 Cytosine Base Editing in Bacteria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 219
Ye Liu, Yang Liu, Ping Zheng, Yu Wang,
and Meng Wang
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233
13.
Contributors
SANGSU BAE •Medical Research Center Organization: Seoul National University College of
Medicine, Seoul, Republic of Korea; Department of Biomedical Sciences, Seoul National
University College of Medicine, Seoul, Republic of Korea; Department of Biochemistry and
Molecular Biology, Seoul National University College of Medicine, Seoul, Republic of Korea
DANIEL E. BAUER • Division of Hematology/Oncology, Boston Children’s Hospital, Boston,
MA, USA; Department of Pediatric Oncology, Dana-Farber Cancer Institute, Boston,
MA, USA; Department of Pediatrics, Harvard Stem Cell Institute, Broad Institute,
Harvard Medical School, Boston, MA, USA
GABRIELE CASIRATI • Division of Hematology/Oncology, Boston Children’s Hospital, Boston,
MA, USA; Department of Pediatric Oncology, Dana-Farber Cancer Institute, Boston,
MA, USA; Department of Pediatrics, Harvard Stem Cell Institute, Broad Institute,
Harvard Medical School, Boston, MA, USA
CHANG SIK CHO • Fight against Angiogenesis-Related Blindness (FARB) Laboratory,
Biomedical Research Institute, Seoul National University Hospital, Seoul, Republic of
Korea
DONGHO CHOI • Department of Surgery, Hanyang University College of Medicine, Seoul,
Republic of Korea; HY Indang Center of Regenerative Medicine and Stem Cell Research,
Hanyang University, Seoul, Republic of Korea; Department of HY-KIST Bio-Convergence,
Hanyang University, Seoul, Republic of Korea
YURI CHOI • Department of Biological Sciences, Korea Advanced Institute for Science and
Technology, Daejeon, Republic of Korea
THOMAS GAJ • Department of Bioengineering, University of Illinois, Urbana, IL, USA; Carl
R. Woese Institute for Genomic Biology, University of Illinois, Urbana, IL, USA
PIETRO GENOVESE • Division of Hematology/Oncology, Boston Children’s Hospital, Boston,
MA, USA; Department of Pediatric Oncology, Dana-Farber Cancer Institute, Boston,
MA, USA; Department of Pediatrics, Harvard Stem Cell Institute, Broad Institute,
Harvard Medical School, Boston, MA, USA
YUTING GUAN • Suzhou Institute of Systems Medicine, Suzhou, China; Shanghai Frontiers
Science Center of Genome Editing and Cell Therapy, Shanghai Key Laboratory of
Regulatory Biology, Institute of Biomedical Sciences and School of Life Sciences, East China
Normal University, Shanghai, China
GUE-HO HWANG • Department of Chemistry, Hanyang University, Seoul, Republic of Korea
HONG-SU JANG • Department of Life Sciences, Korea University, Seoul, Republic of Korea
JAEMIN JEONG • Department of Surgery, Hanyang University College of Medicine, Seoul,
Republic of Korea; HY Indang Center of Regenerative Medicine and Stem Cell Research,
Hanyang University, Seoul, Republic of Korea
TAE YEONG JEONG • Department of Biomedical Sciences and Department of Physiology, Korea
University College of Medicine, Seoul, Republic of Korea
DONG HYUN JO • Department of Anatomy and Cell Biology, Seoul National University
College of Medicine, Seoul, Republic of Korea
ix
14.
x Contributors
MOONYOUNG KANG• Department of Biological Sciences, Korea Advanced Institute for
Science and Technology, Daejeon, Republic of Korea
DAESIK KIM • Department of Precision Medicine, Sungkyunkwan University School of
Medicine, Suwon, Republic of Korea
HUI KWON KIM • Department of Integrative Biotechnology, College of Biotechnology and
Bioengineering, Sungkyunkwan University, Seoul, Republic of Korea
HYEONJIN KIM • Department of Biological Sciences, Korea Advanced Institute for Science
and Technology, Daejeon, Republic of Korea
JEONG HUN KIM • Fight against Angiogenesis-Related Blindness (FARB) Laboratory,
Biomedical Research Institute, Seoul National University Hospital, Seoul, Republic of
Korea; Department of Biomedical Sciences, Seoul National University College of Medicine,
Seoul, Republic of Korea; Department of Ophthalmology, Seoul National University College
of Medicine, Seoul, Republic of Korea; Institute of Reproductive Medicine and Population,
Seoul National University College of Medicine, Seoul, Republic of Korea
KYOUNGMI KIM • Department of Biomedical Sciences and Department of Physiology, Korea
University College of Medicine, Seoul, Republic of Korea
SANG-GYU KIM • Department of Biological Sciences, Korea Advanced Institute for Science
and Technology, Daejeon, Republic of Korea
YOHAN KIM • Max Planck Institute of Molecular Cell Biology and Genetics, Dresden,
Germany
YONGSUB KIM • Department of Biomedical Sciences, Asan Medical Institute of Convergence
Science and Technology, Asan Medical Center, University of Ulsan College of Medicine,
Seoul, Republic of Korea; Stem Cell Immunomodulation Research Center, Asan Medical
Center, University of Ulsan College of Medicine, Seoul, Republic of Korea
SEU-NA LEE • Department of Life Sciences, Korea University, Seoul, Republic of Korea
DALI LI • Suzhou Institute of Systems Medicine, Suzhou, China; Shanghai Frontiers Science
Center of Genome Editing and Cell Therapy, Shanghai Key Laboratory of Regulatory
Biology, Institute of Biomedical Sciences and School of Life Sciences, East China Normal
University, Shanghai, China
COLIN K. W. LIM • Department of Bioengineering, University of Illinois, Urbana, IL, USA
SOO-YEON LIM • Korea Mouse Phenotyping Center, Seoul National University, Seoul,
Republic of Korea
YANG LIU • Key Laboratory of Systems Microbial Biotechnology, Tianjin Institute of
Industrial Biotechnology, Chinese Academy of Sciences, Tianjin, China; National
Technology Innovation Center of Synthetic Biology, Tianjin, China
YE LIU • Key Laboratory of Systems Microbial Biotechnology, Tianjin Institute of Industrial
Biotechnology, Chinese Academy of Sciences, Tianjin, China; National Technology
Innovation Center of Synthetic Biology, Tianjin, China
ANGELO J. MISKALIS • Department of Bioengineering, University of Illinois, Urbana, IL,
USA
MY ANH NGUYEN • Division of Hematology/Oncology, Boston Children’s Hospital, Boston,
MA, USA; Department of Pediatric Oncology, Dana-Farber Cancer Institute, Boston,
MA, USA; Department of Pediatrics, Harvard Stem Cell Institute, Broad Institute,
Harvard Medical School, Boston, MA, USA
YOUNGBIN OH • Department of Biological Sciences, Korea Advanced Institute for Science
and Technology, Daejeon, Republic of Korea
EUNAE PARK • Department of Biological Sciences, Korea Advanced Institute for Science and
Technology, Daejeon, Republic of Korea
15.
Contributors xi
JINMAN PARK• Department of Pharmacology, Yonsei University College of Medicine, Seoul,
Republic of Korea
PABLO PEREZ-PINERA • Department of Bioengineering, University of Illinois, Urbana, IL,
USA; Carl R. Woese Institute for Genomic Biology, University of Illinois, Urbana, IL,
USA; Department of Biomedical and Translational Sciences, Carle-Illinois College of
Medicine, University of Illinois, Urbana, IL, USA; Cancer Center at Illinois, University of
Illinois, Urbana, IL, USA
HIROKI SASAGURI • Laboratory for Proteolytic Neuroscience, RIKEN Center for Brain
Science, Saitama, Japan
JI-EUN SEE • Department of Biomedical Sciences, Asan Medical Institute of Convergence
Science and Technology, Asan Medical Center, University of Ulsan College of Medicine,
Seoul, Republic of Korea; Stem Cell Immunomodulation Research Center, Asan Medical
Center, University of Ulsan College of Medicine, Seoul, Republic of Korea
JE KYUNG SEONG • Korea Mouse Phenotyping Center, Seoul National University, Seoul,
Republic of Korea; Laboratory of Developmental Biology and Genomics, Research Institute
for Veterinary Science, and BK 21 PLUS Program for Creative Veterinary Science
Research, College of Veterinary Medicine, Seoul National University, Seoul, Republic of
Korea; Interdisciplinary Program for Bioinformatics, Seoul National University, Seoul,
Republic of Korea; BIO MAX Institute, Seoul National University, Seoul, Republic of
Korea
BEOMJONG SONG • Medical Research Center Organization: Seoul National University
College of Medicine, Seoul, Republic of Korea
CHUN-QING SONG • Zhejiang University, HangzhouZhejiang, China; School of Life Sciences,
Westlake University, Hangzhou, China; Key Laboratory of Growth Regulation and
Translational Research of Zhejiang Province, School of Life Sciences, Westlake University,
HangzhouZhejiang, China; Westlake Laboratory of Life Sciences and Biomedicine,
HangzhouZhejiang, China
MEIXIA WANG • State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute
of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China
MENG WANG • Key Laboratory of Systems Microbial Biotechnology, Tianjin Institute of
Industrial Biotechnology, Chinese Academy of Sciences, Tianjin, China; National
Technology Innovation Center of Synthetic Biology, Tianjin, China
YU WANG • Key Laboratory of Systems Microbial Biotechnology, Tianjin Institute of
Industrial Biotechnology, Chinese Academy of Sciences, Tianjin, China; National
Technology Innovation Center of Synthetic Biology, Tianjin, China
JAE-SUNG WOO • Department of Life Sciences, Korea University, Seoul, Republic of Korea
JUN WU • Fight against Angiogenesis-Related Blindness (FARB) Laboratory, Biomedical
Research Institute, Seoul National University Hospital, Seoul, Republic of Korea;
Department of Biomedical Sciences, Seoul National University College of Medicine, Seoul,
Republic of Korea
FANG YAN • State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of
Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China
DONG YANG • Zhejiang University, HangzhouZhejiang, China; School of Life Sciences,
Westlake University, Hangzhou, China; Key Laboratory of Growth Regulation and
Translational Research of Zhejiang Province, School of Life Sciences, Westlake University,
HangzhouZhejiang, China; Westlake Laboratory of Life Sciences and Biomedicine,
HangzhouZhejiang, China
16.
xii Contributors
MAN YU• State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of
Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China
JING ZENG • Division of Hematology/Oncology, Boston Children’s Hospital, Boston, MA,
USA; Department of Pediatric Oncology, Dana-Farber Cancer Institute, Boston, MA,
USA; Department of Pediatrics, Harvard Stem Cell Institute, Broad Institute, Harvard
Medical School, Boston, MA, USA
XIAOHUI ZHANG • Institute of Systems Medicine, Chinese Academy of Medical Sciences &
Peking Union Medial College, Beijing, China; Suzhou Institute of Systems Medicine,
Suzhou, China; Shanghai Frontiers Science Center of Genome Editing and Cell Therapy,
Shanghai Key Laboratory of Regulatory Biology, Institute of Biomedical Sciences and School
of Life Sciences, East China Normal University, Shanghai, China
PING ZHENG • Key Laboratory of Systems Microbial Biotechnology, Tianjin Institute of
Industrial Biotechnology, Chinese Academy of Sciences, Tianjin, China; National
Technology Innovation Center of Synthetic Biology, Tianjin, China
HUANBIN ZHOU • State Key Laboratory for Biology of Plant Diseases and Insect Pests,
Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China;
Scientific Observing and Experimental Station of Crop Pests in Guilin, Ministry of
Agriculture and Rural Affairs, Guilin, China
17.
Part I
Overview ofBEs, Diverse Variants of BEs, and Computational
Tools for BEs
4 Beomjong Songand Sangsu Bae
interrupt precise gene editing by inducing the p53-mediated DNA
damage response [7, 8]. Repair of DSB is a key step in gene editing
using conventional CRISPR nucleases, and there are three path-
ways of DSB repair: homology-directed repair (HDR),
non-homologous end joining (NHEJ), and microhomology-
mediated end joining [9–12]. With HDR, precise DNA edits can
be installed by using a donor DNA template, however it is highly
confined to a specific cell cycle phase, S and G2 phases. On the
other hand, operation of the NHEJ and MMEJ pathways is inde-
pendent of cell cycle phases. However, their application is limited
due to error-prone editing results [6, 13].
Alternatively, DNA base editors can edit a gene at a single
nucleotide level with accompanying little DSB generation
[14, 15]. In addition, DNA base editors can be used in both
dividing and non-dividing cells because their operation is not lim-
ited to a specific cell cycle phase. The fact that use of donor
templates is unnecessary also makes DNA base editors even more
of a standout.
2 The Construction of Cytosine Base Editors (CBEs) and the Mechanism of Cytosine
Base Editing
Cytosine base editors (CBEs) commonly contain a fusion form of a
cytidine deaminase and a Cas effector. The initial CBE employed
rAPOBEC1 as a cytidine deaminase and deactivated/dead Cas9
(dCas9) to avoid generation of DSBs (BE1). Recruitment of cyti-
dine deaminase to a target DNA site can occur through an interac-
tion between the dCas9 and the associated single-guide RNA
(sgRNA), with recognition of the single-stranded DNA (ssDNA)
in the R-loop structure formed on the target site. Cytidine deami-
nase then converts deoxycytidines into deoxyuridines. Deoxyuri-
dine preferentially pairs with deoxythymidine, thus the resulting
mismatch U/G pair induces activation of DNA repair pathways
such as base excision repair (BER) and mismatch repair (MMR)
[16]. While the U/G pair can be converted into a T/A pair as
desired through the DNA repair pathways, the U/G pair can be
returned to a C/G pair or non-T/A outcomes can be generated by
the activity of a base excision repair enzyme, uracil N-glycosylase
(UNG) [17]. To reduce such unwanted conversions, uracil glyco-
sylase inhibitor (UGI) peptide was fused to dCas9 (BE2) [14]. In
addition, adoption of Cas9 nickase (nCas9) instead of dCas9
resulted in improved efficiency of base editing by causing the repair
mechanisms to use the strand with deoxyuridine as a template for
repair rather than the opposite strand with a nick (BE3) [14]. For
further suppression of the unwanted conversion, other cytidine
deaminases such as AID (AID-BE3) [18] and PmCDA1 (CDA1-
BE3) [19] were employed instead of rAPOBEC1, or a second UGI
20.
Introduction and Perspectivesof DNA Base Editors 5
was added (BE4) [17]. Improved editing efficiency of BE4 was
achieved through codon optimization (BE4-max) [20] and ances-
tral reconstruction (AncBE4max) [21].
3 The Construction of Adenine Base Editors (ABEs) and the Mechanism of Adenine
Base Editing
The structure and the base editing mechanism of ABEs are similar
to those of CBEs. In ABEs, nCas9 is fused to adenosine deaminase.
All adenosine deaminases discovered so far show activity on RNA
not DNA, therefore, the tRNA-specific adenosine deaminase,
TadA, from Escherichia coli (E. coli) was engineered to operate on
DNA (eTadA*) [15]. ABEs are recruited to the target site of DNA
in the same way as CBEs and convert an adenosine into an inosine,
generating an I/T pair. The mismatched I/T pair is then converted
into a G/C pair through the I/C pair.
The primitive ABE (ABE1.2) comprises the eTadA* monomer.
Because TadAs form a homodimer in nature controlling adenosine
deamination and target binding, respectively, a wild-type TadA
(wtTadA) was fused to eTadA* in the follow-up version of ABE
(ABE2.9). Subsequently, several mutations were introduced into
eTadA* in order to obtain higher editing efficiency in ABE7.9 and
ABE7.10 [15]. Further improvement of ABE7.10 was achieved
through addition of modified NLS and codon optimization in
ABEmax [21]. In the most recent studies, ABE8 variants (ABE8e
and ABE8s) were developed by removing wtTadA and introducing
additional mutations into eTadA* [22, 23].
4 Improvement of DNA Base Editors
4.1 Expanding
Targetable Sites
(Table 1)
Because DNA base editors are based on the CRISPR-Cas system,
targeting DNA sequences is limited by the preference of Cas
nucleases for protospacer adjacent motif (PAM). The original
form of CBEs and ABEs employing Streptococcus pyogenes
(SpCas9) can target only the DNA sequences neighboring the
NGG PAM sequence. Orthologs of Cas effectors having different
PAM preferences were employed, or specific mutations were intro-
duced into the original SpCas9 nucleases in order to expand the
targeting ability of DNA base editors [20, 22–34].
4.2 Modification of
Editing Windows
DNA base editors operate in a confined editing window, which
occasionally results in unwanted base conversion (i.e., bystander
mutation) if multiple Cs or As are located within a editing window.
The frequency of bystander mutations can be reduced by narrowing
the editing window. Several studies have reported that introduction
of mutations in the deaminase determines the width of the editing
window with little effect on the deaminase activity or even with
21.
Deaminase PAM References
6Beomjong Song and Sangsu Bae
Table 1
Diverse combinations of deaminases and Cas effectors
Cas
effectors
Cytidine
deaminase
rAPOBEC1 SpCas9 NGG, NG/GAA/GAT, NGA,
NGAG, NGCG, NG
[14, 17, 21, 24, 32,
35, 47–54]
Spy-mac
Cas9
NAA [20]
SaCas9 NNGRRT, NNNRRT [24]
SauriCas9 NNGG [27]
LbCas12a TTTV [28]
AsCas12a TTTV [29]
ScCas9 NNG [31]
hAPBEC3A spCas9 NGG [18, 36, 50, 52]
PmCDA1 spCas9 NGG, NG [18, 19, 26, 37]
cjCas9 NNNVRYAC [55]
hAID spCas9 NGG [18, 56]
PpAPOBEC1 spCas9 NGG [57]
RrA3F SpCas9 NGG [57]
AmAPOBEC1 SpCas9 NGG [57]
SsAPOBEC3B spCas9 NGG [57]
Adenosine
deaminase
TadA7.10 SpCas9 NGG, NGA, NGCG, NG,NG/
GAA/GAT
[15, 21, 25, 30, 32–
34, 47, 48]
Spy-mac
Cas9
NAA [20]
SaCas9 NNGRRT, NNNRRT [32, 33]
ScCas9 NNG [31]
SauriCas9 NNGG [27]
Nme2Cas9 NNNNCC [58]
TadA8e SpCas9 NGG [22, 23]
SaCas9 NNGRRT [22]
LbCas12a TTTV [22]
AsCas12a TTTV [22]
Cas12f TTTR [46]
TadA8s SpCas9 NGG, NG [23]
SaCas9 NNGRRT [23]
TadA8.17 SpCas9 NGG [23]
TadA7.10 SpCas9 NGG [18, 32, 50, 59]
enhanced editing efficiency [14, 24, 28, 35]. Alternatively, deami-
nases requiring a specific motif can be employed as a method of
lowering the number of bystander mutations [36, 37].
On the other hand, expanding the editing window might be
necessary if there is no Cs or As within the editing window. DNA
base editors having circularly permuted SpCas9 (CP-spCas9)
nucleases have shown broaden editing windows without changes
in PAM preference [32]. In addition, some variants of DNA base
editors with CP-SpCas9 can edit bases upstream of the protospacer
on both target and non-target strands [32].
22.
Introduction and Perspectivesof DNA Base Editors 7
4.3 Generating
Transversion Edits
Conventional DNA base editors are limited to DNA base transition
mutations (purine to purine or pyrimidine to pyrimidine), thus
there is a need for novel DNA base editors that can induce transver-
sion mutations. As mentioned above, inhibition of UNG is
required for C-to-T conversion by CBE. In other words, use of
CBE can result in C/G to non-T/A outcomes through BER path-
ways mediated by UNG. Indeed, removal of UGI from CBEs such
as BE4max resulted in an increase of C-to-G edits compared to the
original BE4max. The bias to C-to-G conversion was enhanced by
addition of UNG protein to CBEs [38]. In addition, C-to-A edit-
ing was induced by replacement of rAPOBEC1 with AID in
prokaryotes [39].
4.4 Generating Dual
Base Conversions of
C-to-T and A-to-G at
Once
Efforts to expand the capabilities of base editing led to the devel-
opment of DNA base editors that can introduce both C-to-T and
A-to-G conversions at the same time. According to several studies
reported around the same time, fusion of both adenosine deami-
nase and cytidine deaminase to a single Cas nuclease exhibits a dual
base editing activity with minimal RNA off-target effects [40].
4.5 TC-Specific Base
Editing by ABE
It was revealed that ABE can introduce unwanted C-to-T edits
within the editing window [41]. For its original purpose, a
D108Q mutation was introduced to the adenosine deaminase to
suppress the cytidine deamination activity. Interestingly, P48R
mutation in the adenosine deaminase showed opposite effect;
cytosine following thymine (i.e., TC) was majorly converted. As a
result, the ABE with P48R mutation could induce TC-to-TT and
TC-to-TG editing with and without UGI, respectively
[42]. Because the cytosine editing by the mutated ABE is promi-
nent at the TC motif, this novel DNA base editor can be used as a
TC-specific base editing tool.
4.6 Adeno-
Associated Virus
(AAV)-Loadable BEs
The availability of efficient and safe delivery methods is a prerequi-
site for use of gene editing tools as therapeutics. Among methods
for virus-based delivery, AAV is widely used due to efficient expres-
sion of its cargo with less toxicity such as the integration of viral
DNA into the host genome. Unfortunately, the size of DNA base
editors, which exceeds the packaging capacity of AAV (~4.7 kb),
causes difficulty in loading DNA base editors into a single AAV
vector. In the effort to bypass this problem, one breakthrough was
achieved by splitting DNA base editors into two parts for packaging
into dual AAV vectors [43, 44]. However, the transduction effi-
ciency of the AAV vector is compromised by the dual AAV system
[45]. Alternatively, successful implantation of a small-sized Cas
nuclease such as Cas12f in ABE was achieved, resulting in genera-
tion of dCasMINI-ABE, which can be loaded into a single
AAV [46].
23.
8 Beomjong Songand Sangsu Bae
5 Conclusions
As if to prove the potential, since they were first showcased, appli-
cation of DNA base editors have shown rapid expansion in various
fields such as biology, biomedical research, and agriculture. Using
DNA base editors, gene editing can be performed with high effi-
ciency and precision, and they are compatible with many cell types.
In addition, use of DNA base editors does not require generation of
DSBs for gene editing, and DNA editing can be performed at a
single nucleotide, further enhancing the prospect for clinical
application.
On the other hand, as with other gene editing tools, there are
some disadvantages associated with use of DNA base editors. DNA
off-target effect, RNA off-target effect, and bystander mutation are
still a possibility when performing gene editing with DNA base
editors. However, findings from an abundance of research have
suggested the use of manipulating Cas nucleases, deaminases, and
guide RNAs methods in order to avoid these problems. Further-
more, the inability to install transversion mutations is regarded as
the most fundamental limitation of DNA base editors, however,
attainment of a deep understanding of biochemical knowledge
regarding DNA repair mechanisms will be helpful in solving this
problem.
Without a doubt, DNA base editors are currently one of the
best genome editing tools, though there are still some problems to
be solved. Efforts to develop improved DNA base editors are
ongoing, and, as a result of these efforts, wider and more preferen-
tial use of DNA base editors will be expected in the future.
Acknowledgments
This research was supported by the National Research Foundation
of Korea (NRF) no. 2021R1I1A1A01056885 to B.S.
References
1. Mali P, Esvelt KM, Church GM (2013) Cas9 as
a versatile tool for engineering biology. Nat
Methods 10(10):957–963. https:/
/doi.org/
10.1038/nmeth.2649
2. Sander JD, Joung JK (2014) CRISPR-Cas sys-
tems for editing, regulating and targeting gen-
omes. Nat Biotechnol 32(4):347–355.
https:/
/doi.org/10.1038/nbt.2842
3. Kim H, Kim JS (2014) A guide to genome
engineering with programmable nucleases.
Nat Rev Genet 15(5):321–334. https:/
/doi.
org/10.1038/nrg3686
4. Doudna JA, Charpentier E (2014) Genome
editing. The new frontier of genome engineer-
ing with CRISPR-Cas9. Science 346(6213):
1258096. https:/
/doi.org/10.1126/science.
1258096
5. Koonin EV, Makarova KS, Zhang F (2017)
Diversity, classification and evolution of
CRISPR-Cas systems. Curr Opin Microbiol
37:67–78. https:/
/doi.org/10.1016/j.mib.
2017.05.008
6. Kosicki M, Tomberg K, Bradley A (2018)
Repair of double-strand breaks induced by
24.
Introduction and Perspectivesof DNA Base Editors 9
CRISPR–Cas9 leads to large deletions and
complex rearrangements. Nat Biotechnol
36(8):765–771. https:/
/doi.org/10.1038/
nbt.4192
7. Haapaniemi E, Botla S, Persson J, Schmierer B,
Taipale J (2018) CRISPR-Cas9 genome edit-
ing induces a p53-mediated DNA damage
response. Nat Med 24(7):927–930. https:/
/
doi.org/10.1038/s41591-018-0049-z
8. Ihry RJ, Worringer KA, Salick MR, Frias E,
Ho D, Theriault K, Kommineni S, Chen J,
Sondey M, Ye C, Randhawa R, Kulkarni T,
Yang Z, McAllister G, Russ C, Reece-Hoyes J,
Forrester W, Hoffman GR, Dolmetsch R, Kay-
kas A (2018) p53 inhibits CRISPR–Cas9 engi-
neering in human pluripotent stem cells. Nat
Med 24(7):939–946. https:/
/doi.org/10.
1038/s41591-018-0050-6
9. Chiruvella KK, Liang Z, Wilson TE (2013)
Repair of double-strand breaks by end joining.
Cold Spring Harb Perspect Biol 5(5):a012757.
https://doi.org/10.1101/cshperspect.
a012757
10. Ma JL, Kim EM, Haber JE, Lee SE (2003)
Yeast Mre11 and Rad1 proteins define a
Ku-independent mechanism to repair double-
strand breaks lacking overlapping end
sequences. Mol Cell Biol 23(23):8820–8828.
https://doi.org/10.1128/mcb.23.23.
8820-8828.2003
11. Moynahan ME, Jasin M (2010) Mitotic
homologous recombination maintains geno-
mic stability and suppresses tumorigenesis.
Nat Rev Mol Cell Biol 11(3):196–207.
https:/
/doi.org/10.1038/nrm2851
12. Lieber MR (2010) The mechanism of double-
strand DNA break repair by the nonhomolo-
gous DNA end-joining pathway. Annu Rev
Biochem 79:181–211. https:/
/doi.org/10.
1146/annurev.biochem.052308.093131
13. Chang HHY, Pannunzio NR, Adachi N, Lie-
ber MR (2017) Non-homologous DNA end
joining and alternative pathways to double-
strand break repair. Nat Rev Mol Cell Biol
18(8):495–506. https:/
/doi.org/10.1038/
nrm.2017.48
14. Komor AC, Kim YB, Packer MS, Zuris JA, Liu
DR (2016) Programmable editing of a target
base in genomic DNA without double-
stranded DNA cleavage. Nature 533(7603):
420–424. https://doi.org/10.1038/
nature17946
15. Gaudelli NM, Komor AC, Rees HA, Packer
MS, Badran AH, Bryson DI, Liu DR (2017)
Programmable base editing of A*T to G*C in
genomic DNA without DNA cleavage. Nature
551(7681):464–471. https:/
/doi.org/10.
1038/nature24644
16. Kunz C, Saito Y, Schar P (2009) DNA repair in
mammalian cells: mismatched repair: variations
on a theme. Cell Mol Life Sci 66(6):
1021–1038. https://doi.org/10.1007/
s00018-009-8739-9
17. Komor AC, Zhao KT, Packer MS, Gaudelli
NM, Waterbury AL, Koblan LW, Kim YB,
Badran AH, Liu DR (2017) Improved base
excision repair inhibition and bacteriophage
Mu Gam protein yields C:G-to-T:A base edi-
tors with higher efficiency and product purity.
Sci Adv 3(8):eaao4774. https:/
/doi.org/10.
1126/sciadv.aao4774
18. Grunewald J, Zhou R, Iyer S, Lareau CA, Gar-
cia SP, Aryee MJ, Joung JK (2019) CRISPR
DNA base editors with reduced RNA off-target
and self-editing activities. Nat Biotechnol
37(9):1041–1048. https:/
/doi.org/10.1038/
s41587-019-0236-6
19. Nishida K, Arazoe T, Yachie N, Banno S,
Kakimoto M, Tabata M, Mochizuki M,
Miyabe A, Araki M, Hara KY, Shimatani Z,
Kondo A (2016) Targeted nucleotide editing
using hybrid prokaryotic and vertebrate adap-
tive immune systems. Science 353(6305).
https:/
/doi.org/10.1126/science.aaf8729
20. Liu Z, Shan H, Chen S, Chen M, Song Y,
Lai L, Li Z (2019) Efficient base editing with
expanded targeting scope using an engineered
Spy-mac Cas9 variant. Cell Discov 5:58.
https://doi.org/10.1038/s41421-019-
0128-4
21. Koblan LW, Doman JL, Wilson C, Levy JM,
Tay T, Newby GA, Maianti JP, Raguram A, Liu
DR (2018) Improving cytidine and adenine
base editors by expression optimization and
ancestral reconstruction. Nat Biotechnol
36(9):843–846. https:/
/doi.org/10.1038/
nbt.4172
22. Richter MF, Zhao KT, Eton E, Lapinaite A,
Newby GA, Thuronyi BW, Wilson C, Koblan
LW, Zeng J, Bauer DE, Doudna JA, Liu DR
(2020) Phage-assisted evolution of an adenine
base editor with improved Cas domain com-
patibility and activity. Nat Biotechnol 38(7):
883–891. https:/
/doi.org/10.1038/s41587-
020-0453-z
23. Gaudelli NM, Lam DK, Rees HA, Sola-Esteves
NM, Barrera LA, Born DA, Edwards A,
Gehrke JM, Lee SJ, Liquori AJ, Murray R,
Packer MS, Rinaldi C, Slaymaker IM, Yen J,
Young LE, Ciaramella G (2020) Directed evo-
lution of adenine base editors with increased
activity and therapeutic application. Nat Bio-
technol 38(7):892–900. https:/
/doi.org/10.
1038/s41587-020-0491-6
24. Kim YB, Komor AC, Levy JM, Packer MS,
Zhao KT, Liu DR (2017) Increasing the
25.
10 Beomjong Songand Sangsu Bae
genome-targeting scope and precision of base
editing with engineered Cas9-cytidine deami-
nase fusions. Nat Biotechnol 35(4):371–376.
https:/
/doi.org/10.1038/nbt.3803
25. Hu JH, Miller SM, Geurts MH, Tang W,
Chen L, Sun N, Zeina CM, Gao X, Rees HA,
Lin Z, Liu DR (2018) Evolved Cas9 variants
with broad PAM compatibility and high DNA
specificity. Nature 556(7699):57–63. https:/
/
doi.org/10.1038/nature26155
26. Nishimasu H, Shi X, Ishiguro S, Gao L,
Hirano S, Okazaki S, Noda T, Abudayyeh
OO, Gootenberg JS, Mori H, Oura S,
Holmes B, Tanaka M, Seki M, Hirano H,
Aburatani H, Ishitani R, Ikawa M, Yachie N,
Zhang F, Nureki O (2018) Engineered
CRISPR-Cas9 nuclease with expanded target-
ing space. Science 361(6408):1259–1262
27. Hu Z, Wang S, Zhang C, Gao N, Li M,
Wang D, Wang D, Liu D, Liu H, Ong SG,
Wang H, Wang Y (2020) A compact Cas9
ortholog from Staphylococcus auricularis
(SauriCas9) expands the DNA targeting
scope. PLoS Biol 18(3):e3000686. https:/
/
doi.org/10.1371/journal.pbio.3000686
28. Li X, Wang Y, Liu Y, Yang B, Wang X, Wei J,
Lu Z, Zhang Y, Wu J, Huang X, Yang L, Chen
J (2018) Base editing with a Cpf1-cytidine
deaminase fusion. Nat Biotechnol 36(4):
324–327. https:/
/doi.org/10.1038/nbt.
4102
29. Kleinstiver BP, Sousa AA, Walton RT, Tak YE,
Hsu JY, Clement K, Welch MM, Horng JE,
Malagon-Lopez J, Scarfo I, Maus MV,
Pinello L, Aryee MJ, Joung JK (2019) Engi-
neered CRISPR-Cas12a variants with increased
activities and improved targeting ranges for
gene, epigenetic and base editing. Nat Biotech-
nol 37(3):276–282. https:/
/doi.org/10.
1038/s41587-018-0011-0
30. Hua K, Tao X, Han P, Wang R, Zhu J-K (2019)
Genome engineering in rice using Cas9 var-
iants that recognize NG PAM sequences. Mol
Plant 12(7):1003–1014. https:/
/doi.org/10.
1016/j.molp.2019.03.009
31. Chatterjee P, Jakimo N, Jacobson JM (2018)
Minimal PAM specificity of a highly similar
Spcas9 ortholog. Science. Advances 4(10):
eaau0766
32. Huang TP, Zhao KT, Miller SM, Gaudelli NM,
Oakes BL, Fellmann C, Savage DF, Liu DR
(2019) Circularly permuted and
PAM-modified Cas9 variants broaden the tar-
geting scope of base editors. Nat Biotechnol
37(6):626–631. https:/
/doi.org/10.1038/
s41587-019-0134-y
33. Yang L, Zhang X, Wang L, Yin S, Zhu B, Xie L,
Duan Q, Hu H, Zheng R, Wei Y, Peng L,
Han H, Zhang J, Qiu W, Geng H, Siwko S,
Zhang X, Liu M, Li D (2018) Increasing tar-
geting scope of adenosine base editors in
mouse and rat embryos through fusion of
TadA deaminase with Cas9 variants. Protein
Cell 9(9):814–819. https:/
/doi.org/10.
1007/s13238-018-0568-x
34. Jeong YK, Yu J, Bae S (2019) Construction of
non-canonical PAM-targeting adenosine base
editors by restriction enzyme-free DNA clon-
ing using CRISPR-Cas9. Sci Rep 9(1):4939.
https://doi.org/10.1038/s41598-019-
41356-1
35. Liu Z, Chen S, Shan H, Jia Y, Chen M, Song Y,
Lai L, Li Z (2020) Efficient base editing with
high precision in rabbits using YFE-BE4max.
Cell Death Dis 11(1):36. https:/
/doi.org/10.
1038/s41419-020-2244-3
36. Gehrke JM, Cervantes O, Clement MK, Wu Y,
Zeng J, Bauer DE, Pinello L, Joung JK (2018)
An APOBEC3A-Cas9 base editor with mini-
mized bystander and off-target activities. Nat
Biotechnol 36(10):977–982. https:/
/doi.org/
10.1038/nbt.4199
37. Tan J, Zhang F, Karcher D, Bock R (2019)
Engineering of high-precision base editors for
site-specific single nucleotide replacement. Nat
Commun 10(1):439. https:/
/doi.org/10.
1038/s41467-018-08034-8
38. Kurt IC, Zhou R, Iyer S, Garcia SP, Miller BR,
Langner LM, Grunewald J, Joung JK (2021)
CRISPR C-to-G base editors for inducing tar-
geted DNA transversions in human cells. Nat
Biotechnol 39(1):41–46. https:/
/doi.org/10.
1038/s41587-020-0609-x
39. Zhao D, Li J, Li S, Xin X, Hu M, Price MA,
Rosser SJ, Bi C, Zhang X (2021) Glycosylase
base editors enable C-to-A and C-to-G base
changes. Nat Biotechnol 39(1):35–40.
https://doi.org/10.1038/s41587-020-
0592-2
40. Sakata RC, Ishiguro S, Mori H, Tanaka M,
Tatsuno K, Ueda H, Yamamoto S, Seki M,
Masuyama N, Nishida K, Nishimasu H,
Arakawa K, Kondo A, Nureki O, Tomita M,
Aburatani H, Yachie N (2020) Base editors for
simultaneous introduction of C-to-T and A-to-
G mutations. Nat Biotechnol 38(7):865–869.
https://doi.org/10.1038/s41587-020-
0509-0
41. Kim HS, Jeong YK, Hur JK, Kim JS, Bae S
(2019) Adenine base editors catalyze cytosine
conversions in human cells. Nat Biotechnol
37(10):1145–1148. https:/
/doi.org/10.
1038/s41587-019-0254-4
42. Jeong YK, Lee S, Hwang GH, Hong SA, Park
SE, Kim JS, Woo JS, Bae S (2021) Adenine
base editor engineering reduces editing of
26.
51.
Introduction and Perspectivesof DNA Base Editors 11
bystander cytosines. Nat Biotechnol 39(11):
1426–1433. https://doi.org/10.1038/
s41587-021-00943-2
43. Levy JM, Yeh WH, Pendse N, Davis JR,
Hennessey E, Butcher R, Koblan LW,
Comander J, Liu Q, Liu DR (2020) Cytosine
and adenine base editing of the brain, liver,
retina, heart and skeletal muscle of mice via
adeno-associated viruses. Nat Biomed Eng
4(1):97–110. https:/
/doi.org/10.1038/
s41551-019-0501-5
44. Ryu SM, Koo T, Kim K, Lim K, Baek G, Kim
ST, Kim HS, Kim DE, Lee H, Chung E, Kim JS
(2018) Adenine base editing in mouse
embryos and an adult mouse model of Duch-
enne muscular dystrophy. Nat Biotechnol
36(6):536–539. https:/
/doi.org/10.1038/
nbt.4148
45. Colella P, Ronzitti G, Mingozzi F (2018)
Emerging issues in AAV-mediated in vivo
gene therapy. Mol Ther Methods Clin Dev 8:
87–104. https:/
/doi.org/10.1016/j.omtm.
2017.11.007
46. Xu X, Chemparathy A, Zeng L, Kempton HR,
Shang S, Nakamura M, Qi LS (2021) Engi-
neered miniature CRISPR-Cas system for
mammalian genome regulation and editing.
Mol Cell 81(20):4333–4345. e4334. https:/
/
doi.org/10.1016/j.molcel.2021.08.008
47. Zuo E, Sun Y, Wei W, Yuan T, Ying W, Sun H,
Yuan L, Steinmetz LM, Li Y, Yang H (2019)
Cytosine base editor generates substantial
off-target single-nucleotide variants in mouse
embryos. Science 364(6437):289–292.
https:/
/doi.org/10.1126/science.aav9973
48. Jin S, Zong Y, Gao Q, Zhu Z, Wang Y, Qin P,
Liang C, Wang D, Qiu J-L, Zhang F, Gao C
(2019) Cytosine, but not adenine, base editors
induce genome-wide off-target mutations in
rice. Science 364(6437):292–296. https:/
/
doi.org/10.1126/science.aaw7166
49. Grunewald J, Zhou R, Garcia SP, Iyer S, Lareau
CA, Aryee MJ, Joung JK (2019)
Transcriptome-wide off-target RNA editing
induced by CRISPR-guided DNA base editors.
Nature 569(7756):433–437. https:/
/doi.org/
10.1038/s41586-019-1161-z
50. Zhou C, Sun Y, Yan R, Liu Y, Zuo E, Gu C,
Han L, Wei Y, Hu X, Zeng R, Li Y, Zhou H,
Guo F, Yang H (2019) Off-target RNA muta-
tion induced by DNA base editing and its elim-
ination by mutagenesis. Nature 571(7764):
275–278. https:/
/doi.org/10.1038/s41586-
019-1314-0
Doman JL, Raguram A, Newby GA, Liu DR
(2020) Evaluation and minimization of Cas9-
independent off-target DNA editing by cyto-
sine base editors. Nat Biotechnol 38(5):
620–628. https:/
/doi.org/10.1038/s41587-
020-0414-6
52. Zuo E, Sun Y, Yuan T, He B, Zhou C, Ying W,
Liu J, Wei W, Zeng R, Li Y, Yang H (2020)
High-fidelity base editor with no detectable
genome-wide off-target effects. Biorxiv.
https://doi.org/10.1101/2020.02.07.
939074
53. Rees HA, Komor AC, Yeh WH, Caetano-
Lopes J, Warman M, Edge ASB, Liu DR
(2017) Improving the DNA specificity and
applicability of base editing through protein
engineering and protein delivery. Nat Com-
mun 8:15790. https:/
/doi.org/10.1038/
ncomms15790
54. Lee JK, Jeong E, Lee J, Jung M, Shin E, Kim
YH, Lee K, Jung I, Kim D, Kim S, Kim JS
(2018) Directed evolution of CRISPR-Cas9
to increase its specificity. Nat Commun 9(1):
3048. https:/
/doi.org/10.1038/s41467-
018-05477-x
55. Nakagawa R, Ishiguro S, Okazaki S, Mori H,
Tanaka M, Aburatani H, Yachie N,
Nishimasu H, Nureki O (2022) Engineered
Campylobacter jejuni Cas9 variant with
enhanced activity and broader targeting
range. Commun Biol 5(1):211. https:/
/doi.
org/10.1038/s42003-022-03149-7
56. Cheng TL, Li S, Yuan B, Wang X, Zhou W, Qiu
Z (2019) Expanding C-T base editing toolkit
with diversified cytidine deaminases. Nat Com-
mun 10(1):3612. https:/
/doi.org/10.1038/
s41467-019-11562-6
57. Yu Y, Leete TC, Born DA, Young L, Barrera
LA, Lee SJ, Rees HA, Ciaramella G, Gaudelli
NM (2020) Cytosine base editors with mini-
mized unguided DNA and RNA off-target
events and high on-target activity. Nat Com-
mun 11(1):2052. https:/
/doi.org/10.1038/
s41467-020-15887-5
58. Zhang H, Bamidele N, Liu P, Ojelabi O, Gao
XD, Rodriguez T, Cheng H, Kelly K, Watts JK,
Xie J, Gao G, Wolfe SA, Xue W, Sontheimer EJ
(2022) Adenine base editing in vivo with a
single adeno-associated virus vector. GEN Bio-
technol 1(3):285–299. https:/
/doi.org/10.
1089/genbio.2022.0015
59. Rees HA, Wilson C, Doman JL, Liu DR
(2019) Analysis and minimization of cellular
RNA editing by DNA adenine base editors.
Science. Advances 5(5):eaax5717. https:/
/
doi.org/10.1126/sciadv.aax5717
14 Gue-Ho Hwangand Sangsu Bae
conversion using cytidine deaminase, and adenine base editors
(ABEs) [2], which make A-to-G conversion using adenosine deam-
inase. C-to-G transversion (C:G to G:C base editors, CGBEs) was
developed in 2021 by Wei Leong Chew’s group using uracil-DNA
glycosylase [10, 11]. Various versions of base editors, such as
BE4max [12], which increases efficiency in mammalian cell types;
CBE-NG [13], which increases coverage of the protospacer adja-
cent motif (PAM) sequence; enAsCas12a-BE [14], which applies
AsCas12a instead of SpCas9; ABE8e [15], which increases the
accuracy of ABEs; and Target-AID [16, 17], which decreases the
range of the editing window, have been developed in an effort to
improve efficiency, accuracy, and window coverage.
Design of target sequences and analysis of NGS data are diffi-
cult without use of computational programs. In addition to the
PAM sequence and termination of the U6 promoter [18], which
are considered when designing the CRISPR-Cas9 target sequence,
the substitution pattern and base editing window according to the
type of base editor are also considered in the design of the target
sequence for base editors. In addition, because Cas9 can recognize
DNA sequences that are similar to the target sequence, off-target
sequences are also considered in the process of design [19, 20].
Therefore, through the adoption of programs for off-target analy-
sis, such as Cas-OFFinder [21] and CROP-IT [22], the program
for use in designing target sequences for base editor, such as
BE-Designer [23], was developed with consideration of off-target
sequences. There are also programs for use in prediction, such as
DeepBaseEditor [24] and BE-Hive [25], which predict the effi-
ciency or outcome of base editors using machine learning prior to
conduct of experiments. In the progress of the analysis, NGS results
should be aligned to a reference sequence and classified according
to their mutation patterns. However, because NGS data include
too many reads, analysis of the data cannot be performed without
use of computational programs. BE-Analyzer [23] and CRIS-
PResso2 [26] were developed for the analysis of NGS results for
base editors.
According to the running environment, there are a variant of
programs that provide only off-line support, that use a web
browser, and that can function under both conditions. Installation
of additional programs or knowledge of the programming language
may be required for use of off-line programs; thus use of off-line
programs is difficult without computational knowledge. Therefore,
the computational programs that use a web browser in design and
analysis of base editor outcome are selectively described in this
manuscript according to the purpose of each program, as follows:
(1) design tool (BE-Designer), (2) efficiency prediction tool
(DeepBaseEditor), (3) outcome prediction tool (BE-Hive), and
(4) analysis tool (BE-analyzer and CRISPResso2).
29.
Web-Based Computational Toolsfor Base Editors 15
2 Materials
2.1 Web Browser Five web tools described in this paper can be used in various web
browsers, such as Chrome, Edge, Firefox, and Safari. It is recom-
mended to update your web browser with the latest version of it.
3 Methods
3.1 Guide RNA
Design Tool: BE-
Designer
When designing BEs, researchers should consider several condi-
tions, including GC contents, the number of off-targets, and
changes in amino acids according to the properties of the base
editors. BE-Designer, a dedicated web tool for base editors, finds
all possible target sequences for BEs under the useful conditions
described above.
1. Enter the BE-Designer website (http:/
/www.rgenome.net/
be-designer/). BE-Designer can also be accessed through the
Rgenome website.
2. Select the PAM sequence and targeted genome (see Notes 1
and 2).
3. Write the sequence for use in design of the base editor target
sequences on the target sequence box (see Note 3).
4. Select the base editing type and adjust the base editing window.
5. Click the submit button.
6. BE-Designer shows all targetable sequences from the received
sequence with information such as GC contents, position,
direction, and expected changes in the amino acid sequence
(Fig. 1). The information regarding off-target is updated later
when the analysis has been completed. By clicking on the row
in the table, BE-Designer directly shows the target region and
expected changes in the amino acid sequence in the received
sequence in the upper figure (see Note 4).
3.2 Efficiency
Prediction Tool:
DeepBaseEditor
DeepBaseEditor is a web tool used for prediction of efficiency for
the target sequence of ABEs and CBEs. DeepBaseEditor was devel-
oped by training a machine learning model on a large sequencing
data set obtained from high-throughput sequencing. DeepBaseE-
ditor finds the targetable sequence of base editors from the received
sequence and scores for each target efficiency.
1. Enter the DeepBaseEditor website (http:/
/deepcrispr.info/
DeepBaseEditor/).
2. Select the model between CBE and ABE.
3. Write the sequence for design of the target sequences for the
base editor (see Note 5).
30.
16 Gue-Ho Hwangand Sangsu Bae
Fig. 1 BE-Designer results page. BE-Designer shows all targetable sequences and information about each
sequence in the table and the expected changes in the amino acid sequence in the figure
Fig. 2 DeepBaseEditor results page. DeepBaseEditor shows the table with targetable sequence and efficiency
score
4. Click the submit button.
5. DeepBaseEditor shows the results in the table with targetable
sequence and efficiency score. The higher the efficiency score
is, the more efficient the sequence is expected to be (Fig. 2)
(see Note 6).
3.3 Outcome
Prediction Tool:
BE-Hive
BE-Hive predicts the outcomes of base editing and editing effi-
ciency for base editors. Like DeepBaseEditor, BE-Hive was devel-
oped by use of machine learning. BE-Hive shows the predicted
outcomes of base editing and predicted efficiency for each outcome
in the combination of various conditions in the base editor and cell
types.
1. Enter the BE-Hive website (https:/
/www.crisprbehive.design/).
2. Select the model for the combination of base editor and cell
types (see Note 7).
3. Write the sequence for design of the base editor target
sequence with at least 20 nt on the 50
side and at least 10 nt
on the 30
side.
4. Select an appropriate CRISPR protospacer from among the
targetable sequences in the model.
31.
Web-Based Computational Toolsfor Base Editors 17
Fig. 3 BE-Hive results page. BE-Hive shows the top ten major predicted outcomes and predicted efficiency
5. BE-Hive directly shows the results on the page shown below.
BE-Hive shows the top ten major outcome patterns with pre-
dicted efficiency for each pattern and shows the graph for
predicted efficiency of base editing (Fig. 3) (see Note 8). The
prediction results can be downloaded as an Excel file.
3.4 Analysis Tool:
BE-Analyzer
BE-Analyzer is a dedicated analysis web tool for base editors.
BE-Analyzer receives the NGS file and shows the editing efficiency
and mutation frequency with interactive graphs. An advantage of
the BE-Analyzer is that NGS files are not required to be uploaded
to the server during the process of analysis using JavaScript.
1. Enter the BE-Analyzer website (http:/
/www.rgenome.net/be-
analyzer). BE-Analyzer can also be accessed through the Rge-
nome website.
2. Select the NGS Fastq file in the “Sequencing Data” box. Con-
trol data is not mandatory.
3. Write the reference sequence and target sequence, and select
the PAM sequence, the type of base editor.
4. Click the submit button (see Note 9).
5. BE-Analyzer shows the table for the count of each mutation
pattern (Fig. 4a) and the table for each nucleotide substitution
frequency for each position (Fig. 4b) (see Note 10) and shows
the graphs for the substitution frequency at each position
(Fig. 4c). BE-Analyzer also shows the alignment results at the
bottom of the results page (Fig. 4d) (see Note 11).
3.5 Analysis Tool:
CRISPResso2
CRISPResso2 is another web tool for analysis of Cas9, Cas12a,
base editors, and prime editors. CRISPResso2 receives NGS Fastq
files and shows various graphs for results of analysis.
1. Enter the CRISPResso2 website (https:/
/crispresso.pinellolab.
partners.org/).
32.
18 Gue-Ho Hwangand Sangsu Bae
Fig. 4 BE-Analyzer results page. (a) The counts for each mutation pattern. (b) Table showing substitution
frequency at each position. (c) Interactive graphs for substitution frequency. (d) Alignment results
2. Click the “Base editors” button in the “Editing tool” menu.
3. Choose the NGS files and write the amplicon and sgRNA
sequence (see Note 12).
4. Click the “Optional parameters,” and select the appropriate
base editor pattern in the “Base editing” section (see Note 13).
5. Click the submit button.
33.
Web-Based Computational Toolsfor Base Editors 19
Fig. 5 CRISPResso2 results page. (a) Pie chart for mutation reads ratio. (b) Nucleotide mutation percentage for
each position. (c) Allele frequency table. (d) Bar plot for substitution frequency for each position. (e) Bar plot for
substitution frequency for each substitution pattern
6. CRISPResso2 shows the mutation frequency by bar and pie
graphs. First, CRISPResso2 shows the results that are in the
same result format when running Cas9 mode, and the results
for base editors are located on the bottom page. CRISPResso2
analyzes the counts of insertion, deletion, and substitutions
and shows the substitution proportion more specifically
(Fig. 5) (see Note 14).
4 Notes
1. 5’-NG-30
and 50
-NRY-30
are selectable. However, due to server
overload, BE-Designer does not perform an analysis of off-
target counts for two PAM sequences.
2. If there is no desired PAM sequence or target genome, send an
e-mail to the server administrator.
3. The sequence can also be entered as a FASTA-formatted file in
BE-Designer. The sequence has a limit of 1 kb in length.
34.
20 Gue-Ho Hwangand Sangsu Bae
4. The results can be downloaded as a text file, and the same result
can be displayed in BE-Designer for up to 3 days through the
URL link on the results page.
5. The sequence can also be entered as a FASTA-formatted and
compressed file by zipping, and it must be longer than 34 bp.
6. The results can be ordered according to each column and can
be downloaded by clicking the “Download Results” button.
7. BE-Hive supports the various options for selecting the model.
However, if there is no appropriate model, read the user guide
page (https:/
/www.crisprbehive.design/guide#celltype).
BE-Hive describes the alternative method when there is no
suitable model.
8. BE-Hive also compares the efficiency of various types of base
editing when entering the batch mode.
9. BE-Analyzer uses a progressing bar to show the progress of the
analysis in real-time.
10. The expected amino acid codon can be shifted using codon
buttons. If the reference sequence is the reverse complemen-
tary sequence, click the check box “Reverse complementary.”
11. Researchers can sort the alignment results for each mutation
pattern using the filter button and can count the reads that
have specific DNA sequences by typing in the “Filtering by
Sequence” box. In addition, the alignment results can be
downloaded as a text file by clicking the “Download Data”
button.
12. CRISPResso2 supports multiple analyses of up to four data
sets. Additional data set input formats can be added by clicking
the plus mark and the blue background button.
13. In optional parameters, there are various options for analysis
except for base editors. Researchers can perform custom ana-
lyses by changing optional parameters.
14. All results and figures can be downloaded as a zipped com-
pressed file by clicking the “Download report” button at the
bottom of the page.
Acknowledgments
This research was supported by the National Research Foundation
of Korea (NRF) no.2021R1A2C3012908 to S.B.
35.
Web-Based Computational Toolsfor Base Editors 21
References
1. Komor AC, Kim YB, Packer MS, Zuris JA, Liu
DR (2016) Programmable editing of a target
base in genomic DNA without double-
stranded DNA cleavage. Nature 533(7603):
420–424. https://doi.org/10.1038/
nature17946
2. Gaudelli NM, Komor AC, Rees HA, Packer
MS, Badran AH, Bryson DI, Liu DR (2017)
Programmable base editing of A*T to G*C in
genomic DNA without DNA cleavage. Nature
551(7681):464–471. https:/
/doi.org/10.
1038/nature24644
3. Doudna JA, Charpentier E (2014) Genome
editing. The new frontier of genome engineer-
ing with CRISPR-Cas9. Science 346(6213):
1258096. https:/
/doi.org/10.1126/science.
1258096
4. Mali P, Esvelt KM, Church GM (2013) Cas9 as
a versatile tool for engineering biology. Nat
Methods 10(10):957–963. https:/
/doi.org/
10.1038/nmeth.2649
5. Kim H, Kim JS (2014) A guide to genome
engineering with programmable nucleases.
Nat Rev Genet 15(5):321–334. https:/
/doi.
org/10.1038/nrg3686
6. Hsu PD, Lander ES, Zhang F (2014) Devel-
opment and applications of CRISPR-Cas9 for
genome engineering. Cell 157(6):1262–1278.
https:/
/doi.org/10.1016/j.cell.2014.05.010
7. Mali P, Yang L, Esvelt KM, Aach J, Guell M,
DiCarlo JE, Norville JE, Church GM (2013)
RNA-guided human genome engineering via
Cas9. Science 339(6121):823–826. https:/
/
doi.org/10.1126/science.1232033
8. Yang H, Wang H, Shivalila CS, Cheng AW,
Shi L, Jaenisch R (2013) One-step generation
of mice carrying reporter and conditional
alleles by CRISPR/Cas-mediated genome
engineering. Cell 154(6):1370–1379.
https:/
/doi.org/10.1016/j.cell.2013.08.022
9. Jeong YK, Song B, Bae S (2020) Current status
and challenges of DNA base editing tools. Mol
Ther 28(9):1938–1952. https:/
/doi.org/10.
1016/j.ymthe.2020.07.021
10. Chen L, Park JE, Paa P, Rajakumar PD, Prekop
HT, Chew YT, Manivannan SN, Chew WL
(2021) Programmable C:G to G:C genome
editing with CRISPR-Cas9-directed base exci-
sion repair proteins. Nat Commun 12(1):
1384. https:/
/doi.org/10.1038/s41467-
021-21559-9
11. Koblan LW, Arbab M, Shen MW, Hussmann
JA, Anzalone AV, Doman JL, Newby GA,
Yang D, Mok B, Replogle JM, Xu A, Sisley
TA, Weissman JS, Adamson B, Liu DR
(2021) Efficient C*G-to-G*C base editors
developed using CRISPRi screens, target-
library analysis, and machine learning. Nat Bio-
technol 39(11):1414–1425. https:/
/doi.org/
10.1038/s41587-021-00938-z
12. Koblan LW, Doman JL, Wilson C, Levy JM,
Tay T, Newby GA, Maianti JP, Raguram A, Liu
DR (2018) Improving cytidine and adenine
base editors by expression optimization and
ancestral reconstruction. Nat Biotechnol
36(9):843–846. https:/
/doi.org/10.1038/
nbt.4172
13. Hua K, Tao X, Han P, Wang R, Zhu JK (2019)
Genome engineering in rice using Cas9 var-
iants that recognize NG PAM sequences. Mol
Plant 12(7):1003–1014. https:/
/doi.org/10.
1016/j.molp.2019.03.009
14. Kleinstiver BP, Sousa AA, Walton RT, Tak YE,
Hsu JY, Clement K, Welch MM, Horng JE,
Malagon-Lopez J, Scarfo I, Maus MV,
Pinello L, Aryee MJ, Joung JK (2019) Engi-
neered CRISPR-Cas12a variants with increased
activities and improved targeting ranges for
gene, epigenetic and base editing. Nat Biotech-
nol 37(3):276–282. https:/
/doi.org/10.
1038/s41587-018-0011-0
15. Richter MF, Zhao KT, Eton E, Lapinaite A,
Newby GA, Thuronyi BW, Wilson C, Koblan
LW, Zeng J, Bauer DE, Doudna JA, Liu DR
(2020) Phage-assisted evolution of an adenine
base editor with improved Cas domain com-
patibility and activity. Nat Biotechnol 38(7):
883–891. https:/
/doi.org/10.1038/s41587-
020-0453-z
16. Grunewald J, Zhou R, Iyer S, Lareau CA, Gar-
cia SP, Aryee MJ, Joung JK (2019) CRISPR
DNA base editors with reduced RNA off-target
and self-editing activities. Nat Biotechnol
37(9):1041–1048. https:/
/doi.org/10.1038/
s41587-019-0236-6
17. Nishida K, Arazoe T, Yachie N, Banno S,
Kakimoto M, Tabata M, Mochizuki M,
Miyabe A, Araki M, Hara KY, Shimatani Z,
Kondo A (2016) Targeted nucleotide editing
using hybrid prokaryotic and vertebrate adap-
tive immune systems. Science 353(6305).
https:/
/doi.org/10.1126/science.aaf8729
18. Gao Z, Herrera-Carrillo E, Berkhout B (2018)
Delineation of the exact transcription termina-
tion signal for type 3 polymerase III. Mol Ther
Nucleic Acids 10:36–44. https:/
/doi.org/10.
1016/j.omtn.2017.11.006
19. Pattanayak V, Lin S, Guilinger JP, Ma E,
Doudna JA, Liu DR (2013) High-throughput
profiling of off-target DNA cleavage reveals
36.
22 Gue-Ho Hwangand Sangsu Bae
RNA-programmed Cas9 nuclease specificity.
Nat Biotechnol 31(9):839–843. https:/
/doi.
org/10.1038/nbt.2673
20. Fu Y, Foden JA, Khayter C, Maeder ML,
Reyon D, Joung JK, Sander JD (2013) High-
frequency off-target mutagenesis induced by
CRISPR-Cas nucleases in human cells. Nat
Biotechnol 31(9):822–826. https:/
/doi.org/
10.1038/nbt.2623
21. Bae S, Park J, Kim JS (2014) Cas-OFFinder: a
fast and versatile algorithm that searches for
potential off-target sites of Cas9 RNA-guided
endonucleases. Bioinformatics 30(10):
1473–1475. https:/
/doi.org/10.1093/bioin
formatics/btu048
22. Singh R, Kuscu C, Quinlan A, Qi Y, Adli M
(2015) Cas9-chromatin binding information
enables more accurate CRISPR off-target pre-
diction. Nucleic Acids Res 43(18):e118.
https:/
/doi.org/10.1093/nar/gkv575
23. Hwang GH, Park J, Lim K, Kim S, Yu J, Yu E,
Kim ST, Eils R, Kim JS, Bae S (2018)
Web-based design and analysis tools for
CRISPR base editing. BMC Bioinform 19(1):
542. https:/
/doi.org/10.1186/s12859-018-
2585-4
24. Song M, Kim HK, Lee S, Kim Y, Seo SY,
Park J, Choi JW, Jang H, Shin JH, Min S,
Quan Z, Kim JH, Kang HC, Yoon S, Kim
HH (2020) Sequence-specific prediction of
the efficiencies of adenine and cytosine base
editors. Nat Biotechnol 38(9):1037–1043.
https://doi.org/10.1038/s41587-020-
0573-5
25. Arbab M, Shen MW, Mok B, Wilson C,
Matuszek Z, Cassa CA, Liu DR (2020) Deter-
minants of base editing outcomes from target
library analysis and machine learning. Cell
182(2):463–480. e430. https:/
/doi.org/10.
1016/j.cell.2020.05.037
26. Clement K, Rees H, Canver MC, Gehrke JM,
Farouni R, Hsu JY, Cole MA, Liu DR, Joung
JK, Bauer DE, Pinello L (2019) CRISPResso2
provides accurate and rapid genome editing
sequence analysis. Nat Biotechnol 37(3):
224–226. https:/
/doi.org/10.1038/s41587-
019-0032-3
24 Jinman Parkand Hui Kwon Kim
and editing outcomes of base editing is of utmost importance albeit
through time-consuming experiments including preparation of
single-guide RNA (sgRNA), delivery of the sgRNA-encoding vec-
tor to target cells together with ABE- or CBE-encoding vector, and
PCR amplification of target DNA region followed by deep
sequencing.
Several computational models have been developed for predict-
ing the CRISPR-Cas9 nuclease-directed genome editing efficien-
cies [3–9] or the outcomes [10, 11]. However, these models show
limited performance in predicting base editor activity even at the
same target DNA sequences. DeepABE and DeepCBE are deep
learning-based computational models that accurately predict the
base editing efficiencies and outcomes of ABE and CBE, respec-
tively, without laborious experiments [12]. DeepABE and
DeepCBE predict the base editing efficiencies and outcome fre-
quencies using the features that affect the genome editing efficien-
cies and outcomes at given target sequences. These models are
provided as web tools available on https:/
/deepcrispr.info/
DeepBaseEditor. Here, we have prepared a step-by-step procedure
in procuring the target sequence information as input for the web
tools (Fig. 1) and for interpreting the prediction results (Fig. 2)
which will greatly facilitate genome editing using ABE and CBE.
Fig. 1 The model selection and sequence input page of DeepBaseEditor
39.
DeepBaseEditor for BaseEditing 25
Fig. 2 Output tables of DeepABE
2 Materials
Prepare a computer with minimum system requirements for web
browsing to predict base editing efficiencies and outcomes on the
DeepBaseEditor website.
3 Methods
1. Select model: As a first step, select the type of base editor to be
used. DeepBaseEditor provides predicted editing efficiencies
and outcome frequencies for both ABE and CBE (Fig. 1a).
2. Input target sequence: Enter the target DNA sequence of inter-
est into the input sequence box (Fig. 1b). The sequence should
be longer than 26 nt because the DeepBaseEditor uses 24~26-
nt-long wide target sequences, which include a 1~3-nt left
neighbor, a 20-nt guide sequence binding region and a 3-nt
protospacer adjacent motif (PAM; 5′-NGG-3′ sequence) for
predicting base editing efficiencies and outcomes (see Notes 1
40.
26 Jinman Parkand Hui Kwon Kim
and 2) [12]. The wide target sequences should be entered in
FASTA format, starting with a description line followed by
lines of target sequence information. The description line
should begin with “>” symbols to distinguish it from the
following lines with the target sequences. Alternatively, a .zip
file which contains FASTA or FA file can be directly uploaded
to the web server. Examples of input sequence for DeepBaseE-
ditor are as follows:
>example1
ATAACACCGATGACATATACGACAAGTGACGGAGGCA
GATAC
>example2
CTGATTAGACAGATATAAACAGTGGCAAACGTAGG
AAATATAA
3. Submission of the input target sequence: Next, the sequence of
interest can be submitted to the server by clicking the “submit”
button on the web page (Fig. 1c).
4. Find all possible guide RNA sequences within the input sequences:
After the sequence submission, DeepBaseEditor automatically
searches the 5′-NGG-3′ PAM sequences and their
corresponding guide RNA sequences on either DNA strand
and shows the output table containing ID, location, target
sequence, guide RNA sequence, and GC contents of guide
RNA sequences (Fig. 2a). The 5′-NGG-3′ PAM sequence for
each target sequence is shown in red. The ID and location
indicate the description of FASTA format input sequence and
the start:end position of the target sequences within the input
sequence, respectively.
5. Prediction of base editing efficiency for the target sequences:
DeepBaseEditor automatically extracts features including
position-independent and position-dependent nucleotides
and dinucleotides, melting temperature, GC counts, and the
minimum self-folding free energy from the target sequences
(see Note 3). Next, it predicts the base editing efficiencies using
the features and shows the predicted base editing efficiencies on
the output table (Fig. 2a). Here, ABE efficiency and CBE
efficiency refer to the predicted frequency of NGS read counts
(or DNA copies) that contain the intended base-edited
sequences (A to G and C to T conversion for ABE and CBE,
respectively) within the wide editable window (positions 3–10;
see Notes 4 and 5). The output is a downloadable .txt file, and
the user can use the URL shown on the result page for later
access.
6. Prediction of all possible outcome sequences and predicted outcome
frequencies for each target sequences: If there are more than one
target adenine or cytosine in the editable window, base editing
41.
DeepBaseEditor for BaseEditing 27
usually generates diverse outcome sequences. When the user
clicks the ID in each row, DeepBaseEditor provides a table with
all possible outcome sequences, relative proportion (ABE pro-
portion and CBE proportion), and the absolute frequency
(DeepABE and DeepCBE scores) of each outcome sequence
(Fig. 2b; see Notes 6 and 7). The output is a downloadable .txt
file, and the user can use the URL shown on the result page for
later access.
7. Interpretation of prediction scores: DeepABE and DeepCBE
accurately predict the efficiencies and outcome frequencies of
ABE- and CBE-mediated editing at given target sequences
indicated with Pearson’s correlations ranging from 0.50 to
0.95 in HEK293T, HCT116, and U2OS cell lines and
human induced pluripotent stem cells (iPSCs). However, the
absolute editing frequencies of ABE and CBE can vary depend-
ing on the experimental conditions including the cell type,
delivery efficiency, incubation time, and the version of deami-
nase domain (see Notes 8 and 9) [12].
8. Batch run: If there are thousands of target sequences to be
tested, the user may download and use the source codes for
DeepABE and DeepCBE available on Github (https:/
/github.
com/MyungjaeSong/Paired-Library/tree/DeepCRISPR.
info/DeepBaseEditor). The required versions of Python and
various dependent packages are listed below and also available
on the Github page. Briefly, the latest successful run was con-
ducted on Python (2.7.12) using numpy (1.14.5), scipy
(1.1.0), Tensorflow (1.4.1) with CUDA (8.0.61), and
cuDNN (6.0.21). The input target sequences can be listed on
a .txt file using the file format:>example ABE_Efficiency
input1: ./ABE_Efficiency_sample.txt # List of Target
Sequence(s)
Target number 30 bp target sequence (4 bp + 20 bp proto-
spacer + PAM + 3 bp) 1 AACTGAAGGCTGAACAGCA
GGGGTGGGAGA
input2: ./ABE_Efficiency_Weight/ # Pre-trained Weight Files
The input target sequence and pre-trained weight files
should be located in the same working directory as the main
script file, TEST_ABE_Efficiency.py. The user can make modi-
fication to the system paths and parameters by editing the
sections labeled “## System Paths ##” and “## Run Para-
meters ##” within the source code, respectively.
The batch run procedure for CBE is as described above for
ABE with the respective input and script files.>example
CBE_Efficiency
input1: ./CBE_Efficiency_sample.txt # List of Target
Sequence(s)
42.
28 Jinman Parkand Hui Kwon Kim
Target number 30 bp target sequence (4 bp + 20 bp proto-
spacer + PAM + 3 bp) 1 AACTGAAGGCTGAACAGCA
GGGGTGGGAGA
input2: ./CBE_Efficiency_Weight/ # Pre-trained Weight Files
The output file will be generated in a directory named
“output” within the same working directory unless otherwise
modified by the user with the expected file format as below:
>example output
outputs/TEST_OUTPUT_for_test.xlsx
Predicted activity score for sequence 1 : 25.79517365
Following the same principle, detailed outline of the batch run
operation for additional analyses is available on the Github page,
https:/
/github.com/MyungjaeSong/Paired-Library/tree/
DeepCRISPR.info/DeepBaseEditor
4 Notes
1. The optimal length of wide target sequences for DeepABE and
DeepCBE was determined based on the results of cross-
validation. Using 25-nt (2-nt left neighbor + 20-nt guide
sequence binding region + 3-nt PAM sequence) and 24-nt
(1-nt left neighbor + 20-nt guide sequence binding region +
3-nt PAM sequence) wide target sequences showed the highest
performances for the prediction of ABE- and CBE-directed
base editing efficiencies, respectively. The models that use
26-nt (3-nt left neighbor + 20-nt guide sequence binding
region + 3-nt PAM sequence) and 25-nt (2-nt left neighbor +
20-nt guide sequence binding region + 3-nt PAM sequence)
wide target sequences show the highest performances for the
prediction ABE- and CBE-directed base editing outcome pro-
portions, respectively.
2. DeeABE and DeeCBE were developed using data sets of
ABE7.10 (variant of ABE) and BE4 (variant of CBE) mediated
base editing efficiencies and outcome proportions at 13,504
and 14,157 wide target sequences, respectively. ABE7.10 [1]
and BE4 [13] share the Cas9 nickase domain derived from
Streptococcus pyogenes which recognized 5′-NGG-3′ PAM
sequences.
3. Analysis of large-scale data sets allowed for the characterization
of features affecting the ABE and CBE efficiencies and
outcome proportions. These features include position-
independent and position-dependent nucleotides and
43.
Total reads ofa
transitions A T to G
ð Þ
within
R
Total rea
transitions A
ð Þ
w Þ
DeepBaseEditor for Base Editing 29
dinucleotides, melting temperature, GC counts, and the mini-
mum self-folding free energy from the target sequences. Previ-
ous studies have demonstrated that SpCas9 activities were
slightly higher at target sites with high chromatin accessibility
[6]. Comparing DNase I hypersensitive (DHS) sites to
non-DHS sites indicated CBE efficiency was significantly (1.9
fold, P = 0.0077) higher while ABE was less so (1.1 fold,
P = 0.55).
4. We calculated the base editing efficiencies as follows:
Base editing efficiency %
ð Þ =
ll sequences that contain inteanded target nucleotide
C base pairs for ABE : C G to T A base pairs for CBE
the wide editable window at positions 3–10
ð Þ
Total reads
× 100
5. The high-throughput evaluation of base editing efficiencies at
thousands of target sequences showed that the editable win-
dow of ABE (ABE7.10) and CBE (BE4) spans positions 3–10,
although a relatively higher level of base editing can be achieved
at positions 4–8 for both ABE and CBE. Here, position 20 indi-
cates the nucleotide located immediately 5′ upstream of the
PAM sequence.
6. We calculated the base editing outcome proportion as follows:
Base editing outcome proportion =
eads of the specific base‐edited outcome sequence
ds of all sequences that contain intended target nucleotide
T to G C base pairs for ABE; C G to T A base pairs for CBE
ithin the wide editable window at positions 3–10
ð
7. DeepABE and DeepCBE scores indicate the predicted absolute
frequency of each base-edited outcome sequence. Accordingly,
multiplying the base editing efficiency and the base editing
outcome proportion yields the absolute frequency of an out-
come sequence among the total reads (DNA copies). Dee-
pABE and DeepCBE scores are calculated by multiplying the
ABE_proportion and ABE_efficiency or CBE_proportion and
CBE efficiency, respectively.
44.
Absolute frequency ofa
=
Reads of the specific
T
=
Reads
Total reads of all
Transitions A T to
ð Þ
Within the w
×
Total reads of all s
Transitions A T to
ð Þ
Within the w
= base editing outcome
30 Jinman Park and Hui Kwon Kim
n outcome of base‐editing %
ð Þ
base‐edited outcome sequence
otal reads
× %
ð Þ
of the specific base‐edited outcome sequence
sequences that contain intended target nucleotide
G C base pairs for ABE; C G to T A base pairs for CBE
ide editable window at positions 3–10
ð Þ
equences that contain intended target nucleotide
G C base pairs for ABE; C G to T A base pairs for CBE
ide editable window at positions 3–10
ð Þ
Total reads
× 100
proportional × base diting efficiency %
ð Þ
8. Training data sets of DeepABE and DeepCBE were obtained
using the high-throughput evaluation experiments in
HEK293T cells [12]. For the high-throughput evaluation of
base editing activities, a lentiviral library of thousands of guide
RNA-encoding and target sequence pairs that were used previ-
ously for the high-throughput evaluation of Cas9 activities [6]
were adopted. Next, HEK293T cells were transduced with
lentiviral library selected using antibiotics. The cell library was
transiently transfected with either ABE7.10- or BE4-encoding
plasmids. Five days post transfection, cells were harvested and
subjected to deep sequencing to measure the base editing
efficiencies and outcomes at the lentivirally integrated target
sequence. Thus, prediction values of DeepABE and DeepCBE
reflect the absolute editing frequencies of ABE7.10- and
BE4-directed base editing efficiencies measured in HEK293T
cells at 5 days post transfection.
9. Recently, various improved versions of ABE and CBE, involv-
ing biochemical modifications to the deaminase domain, have
been reported [14–20]. However, the Cas9 nickase domain
remains the main catalytic component among these variants.
As such, DeepABE and DeepCBE are expected to accurately
predict the base editing outcomes and efficiencies for these
variants albeit future works can address the potential changes
to the base editing activities elicited by the deaminase domain
modifications.
45.
DeepBaseEditor for BaseEditing 31
References
1. Gaudelli NM, Komor AC, Rees HA, Packer
MS, Badran AH, Bryson DI, Liu DR (2017)
Programmable base editing of A*T to G*C in
genomic DNA without DNA cleavage. Nature
551(7681):464–471. https:/
/doi.org/10.
1038/nature24644
2. Komor AC, Kim YB, Packer MS, Zuris JA, Liu
DR (2016) Programmable editing of a target
base in genomic DNA without
double-stranded DNA cleavage. Nature
533(7603):420–424. https:/
/doi.org/10.
1038/nature17946
3. Chari R, Mali P, Moosburner M, Church GM
(2015) Unraveling CRISPR-Cas9 genome
engineering parameters via a library-on-library
approach. Nat Methods 12(9):823–826.
https:/
/doi.org/10.1038/nmeth.3473
4. Doench JG, Fusi N, Sullender M, Hegde M,
Vaimberg EW, Donovan KF, Smith I,
Tothova Z, Wilen C, Orchard R, Virgin HW,
Listgarten J, Root DE (2016) Optimized
sgRNA design to maximize activity and mini-
mize off-target effects of CRISPR-Cas9. Nat
Biotechnol 34(2):184–191. https:/
/doi.org/
10.1038/nbt.3437
5. Doench JG, Hartenian E, Graham DB,
Tothova Z, Hegde M, Smith I, Sullender M,
Ebert BL, Xavier RJ, Root DE (2014) Rational
design of highly active sgRNAs for CRISPR-
Cas9-mediated gene inactivation. Nat Biotech-
nol 32(12):1262–1267. https:/
/doi.org/10.
1038/nbt.3026
6. Kim HK, Kim Y, Lee S, Min S, Bae JY, Choi
JW, Park J, Jung D, Yoon S, Kim HH (2019)
SpCas9 activity prediction by DeepSpCas9, a
deep learning-based model with high generali-
zation performance. Sci Adv 5(11):eaax9249.
https:/
/doi.org/10.1038/nbt.3026
7. Moreno-Mateos MA, Vejnar CE, Beaudoin
JD, Fernandez JP, Mis EK, Khokha MK, Gir-
aldez AJ (2015) CRISPRscan: designing highly
efficient sgRNAs for CRISPR-Cas9 targeting
in vivo. Nat Methods 12(10):982–988.
https:/
/doi.org/10.1038/nmeth.3543
8. Wong N, Liu W, Wang X (2015)
WU-CRISPR: characteristics of functional
guide RNAs for the CRISPR/Cas9 system.
Genome Biol 16:218. https:/
/doi.org/10.
1186/s13059-015-0784-0
9. Xu H, Xiao T, Chen CH, Li W, Meyer CA,
Wu Q, Wu D, Cong L, Zhang F, Liu JS,
Brown M, Liu XS (2015) Sequence determi-
nants of improved CRISPR sgRNA design.
Genome Res 25(8):1147–1157. https:/
/doi.
org/10.1101/gr.191452.115
10. Bae S, Kweon J, Kim HS, Kim JS (2014)
Microhomology-based choice of Cas9 nuclease
target sites. Nat Methods 11(7):705–706.
https:/
/doi.org/10.1038/nmeth.3015
11. Shen MW, Arbab M, Hsu JY, Worstell D, Cul-
bertson SJ, Krabbe O, Cassa CA, Liu DR, Gif-
ford DK, Sherwood RI (2018) Predictable and
precise template-free CRISPR editing of path-
ogenic variants. Nature 563(7733):646–651.
https://doi.org/10.1038/s41586-018-
0686-x
12. Song M, Kim HK, Lee S, Kim Y, Seo SY,
Park J, Choi JW, Jang H, Shin JH, Min S,
Quan Z, Kim JH, Kang HC, Yoon S, Kim
HH (2020) Sequence-specific prediction of
the efficiencies of adenine and cytosine base
editors. Nat Biotechnol 38(9):1037–1043.
https://doi.org/10.1038/s41587-020-
0573-5
13. Komor AC, Zhao KT, Packer MS, Gaudelli
NM, Waterbury AL, Koblan LW, Kim YB,
Badran AH, Liu DR (2017) Improved base
excision repair inhibition and bacteriophage
Mu Gam protein yields C:G-to-T:A base edi-
tors with higher efficiency and product purity.
Sci Adv 3(8):eaao4774. https:/
/doi.org/10.
1126/sciadv.aao4774
14. Fu J, Li Q, Liu X, Tu T, Lv X, Yin X, Lv J,
Song Z, Qu J, Zhang J, Li J, Gu F (2021)
Human cell based directed evolution of ade-
nine base editors with improved efficiency. Nat
Commun 12(1):5897. https:/
/doi.org/10.
1038/s41467-021-26211-0
15. Gaudelli NM, Lam DK, Rees HA, Sola-Esteves
NM, Barrera LA, Born DA, Edwards A,
Gehrke JM, Lee SJ, Liquori AJ, Murray R,
Packer MS, Rinaldi C, Slaymaker IM, Yen J,
Young LE, Ciaramella G (2020) Directed evo-
lution of adenine base editors with increased
activity and therapeutic application. Nat Bio-
technol 38(7):892–900. https:/
/doi.org/10.
1038/s41587-020-0491-6
16. Gehrke JM, Cervantes O, Clement MK, Wu Y,
Zeng J, Bauer DE, Pinello L, Joung JK (2018)
An APOBEC3A-Cas9 base editor with mini-
mized bystander and off-target activities. Nat
Biotechnol 36(10):977–982. https:/
/doi.org/
10.1038/nbt.4199
17. Grunewald J, Zhou R, Iyer S, Lareau CA, Gar-
cia SP, Aryee MJ, Joung JK (2019) CRISPR
DNA base editors with reduced RNA off-target
and self-editing activities. Nat Biotechnol
37(9):1041–1048. https:/
/doi.org/10.1038/
s41587-019-0236-6
46.
32 Jinman Parkand Hui Kwon Kim
18. Koblan LW, Doman JL, Wilson C, Levy JM,
Tay T, Newby GA, Maianti JP, Raguram A, Liu
DR (2018) Improving cytidine and adenine
base editors by expression optimization and
ancestral reconstruction. Nat Biotechnol
36(9):843–846. https:/
/doi.org/10.1038/
nbt.4172
19. Richter MF, Zhao KT, Eton E, Lapinaite A,
Newby GA, Thuronyi BW, Wilson C, Koblan
LW, Zeng J, Bauer DE, Doudna JA, Liu DR
(2020) Phage-assisted evolution of an adenine
base editor with improved Cas domain com-
patibility and activity. Nat Biotechnol 38(7):
883–891. https:/
/doi.org/10.1038/s41587-
020-0453-z
20. Zuo E, Sun Y, Yuan T, He B, Zhou C, Ying W,
Liu J, Wei W, Zeng R, Li Y, Yang H (2020) A
rationally engineered cytosine base editor
retains high on-target activity while reducing
both DNA and RNA off-target effects. Nat
Methods 17(6):600–604. https:/
/doi.org/
10.1038/s41592-020-0832-x
34 Daesik Kim
targetsite. To define genome-wide specificity of dLbCpf1-BE
without bias, we developed a modified Digenome-seq, which
could capture single-strand breaks (SSBs) [9]. In this method,
purified genomic DNA (gDNA) is incubated with dLbCpf1-BE
in vitro to induce C-to-U conversion at the on-target and potential
off-target sites, and then uracil is removed using uracil DNA gly-
cosylase (UDG) and DNA glycosylase-lyase endonuclease forming
SSBs in the on-/off-target sites. Digested DNA is fragmented,
end-repaired, and ligated to sequencing adaptors to generate
sequencing libraries. The sequencing libraries are subjected to
whole-genome sequencing (WGS), and sequencing reads are
aligned to a reference genome such as hg19 (Human) and mm10
(Mouse). To profile genome-wide off-target sites of dLbCpf1-BE,
we developed a computer program that could find genome-wide
SSBs without bias [9]. Here, we present a detailed experimental
procedure for profiling the genome-wide specificity of
dLbCpf1-BE.
2 Materials
2.1 crRNA DNA
Templates
1. crRNA_F (Macrogen): 5′ - GAA ATT AAT ACG ACT CAC
TAT AAA TTT CTA CTA AGT GTA GAA T -3′.
2. crRNA_R (Macrogen): 5′ - NNN NNN NNN NNN NNN
NNN NNA TTC TAC ACT TAG TAG AAA TTT ATA G -3′
(see Note 1).
3. Phusion High-Fidelity DNA Polymerase (Thermo Scientific).
4. 5× Phusion HF Buffer (Thermo Scientific).
5. dNTPs.
6. TOPcloner™ TA-Blunt Kit (Enzynomics).
7. Exprep Plasmid SV (GeneAll).
8. EcoRI-HF (New England Biolabs).
9. NTPs (ATP, UTP, CTP, and GTP).
10. 100 mM MgCl2.
11. T7 RNA polymerase (New England Biolabs).
12. 1× RNAPol reaction buffer (New England Biolabs): 400 mM
Tris–HCl, 60 mM MgCl2, 10 mM DTT, 20 mM spermidine.
13. RNase inhibitor.
14. 1 M DTT.
15. DNase I (New England Biolabs).
16. 10× DNase I reaction buffer (New England Biolabs): 100 mM
Tris–HCl, 25 mM MgCl2, and 5 mM CaCl2.
17. PCR Product Purification Kit (MGmed).
49.
Co
CBE Profiling UsingDigenome-Seq 35
2.2 Genomic DNA
Extraction
1. DNeasy Blood and Tissue Kit (Qiagen).
2.3 In Vitro Digestion
of Genomic DNA
1. dLbCpf1-BE protein.
2. RNase A.
3. 10× NEB3.1 (New England Biolabs): 100 mM NaCl, 50 mM
Tris–HCl, 10 mM MgCl2, and 100 μg/mL BSA.
4. USER®
Enzyme (NEB).
2.4 Whole-Genome
Sequencing
1. Covaris systems M220 Focused-ultrasonicator.
2. TruSeq DNA PCR-Free Library Kit (Illumina).
3. Agilent Technologies 2100 Bioanalyzer (Agilent).
4. DNA 1000 Chip Kit (Agilent).
5. HiSeq X Ten (Illumina).
6. End Repair Mix (Illumina).
2.5 Digenome-Seq
Analysis
1. SAMtools (http:/
/www.htslib.org/).
3 Methods
3.1 Preparation of
the crRNA
1. Resuspend two complementary oligonucleotides (crRNA_F
and crRNA_R) at a final concentration of 1 μM in the reaction
buffer to get the DNA template for in vitro transcription of
crRNA:
mponent
Volume
(μL)
Final
concentration
NA_F (100 μM) 0.5 1 μM
NA_R (100 μM) 0.5 1 μM
Phusion HF buffer 10 1×
mM dNTPs 2.5 0.5 mM
usion high-Fidelity DNA Polymerase
2 U/μL)
0.5 0.02 U/ μL
clease-free water 36
tal 50
crR
crR
5×
10
Ph
(
Nu
To
2. In a thermocycler, anneal and extend oligonucleotides using
the following program:
50.
C 15 s25
C 15 s
C 15 s
C 1
36 Daesik Kim
Step Temperature Time Cycles
Denaturation 95 °C 1 min 1
Denaturation 95 °
Annealing 56 °
Extension 72 °
Extension 72 °C 3 min 1
Hold 12 °
3. Purify the DNA template using an MG PCR Product Purifica-
tion Kit, following the manufacturer’s instructions.
4. Clone the crRNA DNA template to the pTOP TA-Blunt V2
vector (used for cloning blunt-ended DNA templates) as fol-
lows to reduce oligonucleotide-mediated errors.
mponent Volume (μL)
TOPcloner™ buffer 1
OP TA-Blunt V2 (10 ng/μL) 0.5
A template (5 ng/μL) 1
W. 3.5
tal 6
Co
6×
pT
DN
D.
To
5. After 5 min at room temperature, transform the ligated DNA
into DH5 chemically competent E. coli.
6. Isolate plasmid DNA from transformed colonies using a plas-
mid DNA prep kit, and perform the Sanger sequencing using
M13F primer to validate that the desired sequence has been
inserted into plasmid DNA.
7. By restriction enzyme digestion, linearize plasmids including
the desired DNA template for the crRNA.
mponent Volume (μL)
smid DNA 10 μg (X μL)
oRI-HF 2
t smart 10
W. (88-X)
tal 100
Co
Pla
Ec
Cu
D.
To
8. Incubate the reaction for 4 h at 37 °C, and clean it up with an
MG PCR Product Purification Kit to remove the restriction
enzyme.
51.
mM
CBE Profiling UsingDigenome-Seq 37
9. To make the in vitro transcribed crRNA, mix the DNA tem-
plate for crRNA with T7 RNA polymerase as follows (see
Note 2):
mponent Volume (μL) Final concentration
NA template DNA 600 ng (X μL)
P (100 mM) 2 4 mM
P (100 mM) 2 4 mM
P (100 mM) 2 4 mM
P (100 mM) 2 4 mM
0 mM MgCl2 7 14
RNA polymerase 7.5
ase inhibitor 1.25
DTT 0.5 10 mM
× RNAPol reaction buffer 5 1×
clease-free water (20.75-X)
tal 50
Co
crR
AT
UT
CT
GT
10
T7
RN
1 M
10
Nu
To
10. Perform the in vitro transcription for 4 h at 37 °C, and clean
the in vitro generated crRNA with an MG PCR Product Puri-
fication Kit, following the manufacturer’s instructions,
11. To remove the DNA template used in the in vitro transcription
reaction, incubate the in vitro transcribed crRNA for 30 min
with 1× DNase I reaction buffer and DNase I (final concentra-
tion, 2 U/50 μL). After DNase I treatment, clean up the
mixture once again to remove DNase I using an MG PCR
Product Purification Kit.
3.2 In Vitro
Deamination of
Genomic DNA
1. Prepare genomic DNA from cells using a DNeasy Blood and
Tissue Kit, according to the manufacturer’s instructions.
2. Set up the following process, and incubate it at 37 °C for 8 h to
induce targeted in vitro deamination of genomic DNA, which
induces cytosine to uracil conversion (Fig. 1) (see Note 3).
mponent Volume (μL) Final concentration
nomic DNA 10 μg (X μL)
NA Y 300 nM
bCpf1-BE Z 100 nM
× NEB3.1 50
clease-free water (450-X-Y-Z)
tal 500
Co
Ge
crR
dL
10
Nu
To
52.
38 Daesik Kim
Fig.1 A Digestome-seq method for identifying dLbCpf1-BE off-target sites
3. Add RNase A (final concentration, 50 g/ml), and incubate the
mixture at 25 °C for 10 min to remove the crRNA. Purify the
in vitro digested DNA using a DNeasy Blood and Tissue Kit,
following the manufacturer’s instructions.
4. Amplify the dLbCpf1-BE targeted genomic locus using PCR,
and confirm the cytosine to thymine conversion at the
on-target site using Sanger sequencing (see Note 4).
5. To achieve a uracil excision reaction, incubate the following
mixture for 2 h at 37 °C, and purify the genomic DNA using a
DNeasy Blood and Tissue Kit according to the manufacturer’s
instructions.
mponent Volume (μL) Final concentration
acil-containing genomic DNA
rom step 3
2 μg (X μL)
ER enzyme 6 6 U
× CutSmart buffer 10
clease-free water (84-X)
tal 100
Co
Ur
f
US
10
Nu
To
6. Genomic locus including dLbCpf1-BE target site is amplified
using PCR and confirms the uracil removal at the target site by
Sanger sequencing.
53.
CBE Profiling UsingDigenome-Seq 39
3.3 Digenome-
Sequencing to Profile
dLbCpf1-BE and USER
Mediated Genome-
Wide SSBs
1. Fragment 1 μg of genomic DNA with a Covaris systems ultra-
sonicator using the following parameters to generate a ~350 bp
DNA fragments
ty factor (%) 20
ak/displayed power (W) 50
cles/burst 200
ration (s) 65
mperature (°C) 20
Du
Pe
Cy
Du
Te
2. To generate blunt ends, incubate the fragmented DNA using
End Repair Mix. Ligate the blunt-ended fragments with adap-
tors to create sequencing libraries using an Illumina TruSeq
DNA PCR-Free Library Kit, according to the manufacturer’s
instructions (Fig. 1).
3. Using an Agilent Technologies 2100 Bioanalyzer and a DNA
1000 chip, check the sequencing library quality and size distri-
bution according to the manufacturer’s instructions.
4. Subject sequencing libraries to WGS using an Illumina HiSeq X
Ten Sequencer, following the manufacturer’s instructions (see
Note 5).
5. Map the sequencing reads to the reference genome using Isaac
Aligner with the following parameters.
rameter Value
ase-quality-cutoff 15
eep-duplicates 1
efault-adapters AGATCGGAAGAGC*,*GCTCTTCCGATCT
Pa
--b
--k
--d
3.4 Digenome-Seq
Analysis
1. Using SAMtools, separate the sequence reads into forward and
reverse strands.
Download and install the Digenome-seq program, available at
https:/
/github.com/snugel/
digenome-toolkit (see Note 6).
2. To analyze the BAM file, run the script as follows:
digenome-run -u 0 BAM_file_PATH
54.
40 Daesik Kim
4Notes
1. “NNNNNNNNNNNNNNNNNNNN” represents the target
sequence of LbCpf1-BE, which does not include the PAM
sequence.
2. 10 mM DTT enhances the efficiency of in vitro transcription.
3. Low quality of dLbCpf1-BE protein could induce genomic
DNA degradation.
4. Deamination of cytosine converts it to uracil, which is amplified
to thymine by PCR.
5. The WGS depth should be at least 30× for Digenome-seq
analysis.
6. Alternatively, the Digenome-seq web tool is available at http:/
/
www.rgenome.net/digenome-js.
References
1. Komor AC, Kim YB, Packer MS, Zuris JA, Liu
DR (2016) Programmable editing of a target
base in genomic DNA without double-stranded
DNA cleavage. Nature 533(7603):420–424.
https:/
/doi.org/10.1038/nature17946
2. Nishida K, Arazoe T, Yachie N, Banno S,
Kakimoto M, Tabata M, Mochizuki M,
Miyabe A, Araki M, Hara KY, Shimatani Z,
Kondo A (2016) Targeted nucleotide editing
using hybrid prokaryotic and vertebrate adaptive
immune systems. Science 353(6305). https:/
/
doi.org/10.1126/science.aaf8729
3. Gaudelli NM, Komor AC, Rees HA, Packer MS,
Badran AH, Bryson DI, Liu DR (2017) Pro-
grammable base editing of A*T to G*C in geno-
mic DNA without DNA cleavage. Nature
551(7681):464–471. https:/
/doi.org/10.
1038/nature24644
4. Li X, Wang Y, Liu Y, Yang B, Wang X, Wei J,
Lu Z, Zhang Y, Wu J, Huang X, Yang L, Chen J
(2018) Base editing with a Cpf1-cytidine deam-
inase fusion. Nat Biotechnol 36(4):324–327.
https:/
/doi.org/10.1038/nbt.4102
5. Richter MF, Zhao KT, Eton E, Lapinaite A,
Newby GA, Thuronyi BW, Wilson C, Koblan
LW, Zeng J, Bauer DE, Doudna JA, Liu DR
(2020) Phage-assisted evolution of an adenine
base editor with improved Cas domain compati-
bility and activity. Nat Biotechnol 38(7):
883–891. https:/
/doi.org/10.1038/s41587-
020-0453-z
6. Ryu SM, Koo T, Kim K, Lim K, Baek G, Kim ST,
Kim HS, Kim DE, Lee H, Chung E, Kim JS
(2018) Adenine base editing in mouse embryos
and an adult mouse model of Duchenne muscu-
lar dystrophy. Nat Biotechnol 36(6):536–539.
https:/
/doi.org/10.1038/nbt.4148
7. Zafra MP, Schatoff EM, Katti A, Foronda M,
Breinig M, Schweitzer AY, Simon A, Han T,
Goswami S, Montgomery E, Thibado J, Kasten-
huber ER, Sanchez-Rivera FJ, Shi J, Vakoc CR,
Lowe SW, Tschaharganeh DF, Dow LE (2018)
Optimized base editors enable efficient editing
in cells, organoids and mice. Nat Biotechnol
36(9):888–893. https:/
/doi.org/10.1038/
nbt.4194
8. Yeh WH, Chiang H, Rees HA, Edge ASB, Liu
DR (2018) In vivo base editing of post-mitotic
sensory cells. Nat Commun 9(1):2184. https:/
/
doi.org/10.1038/s41467-018-04580-3
9. Kim D, Lim K, Kim DE, Kim JS (2020)
Genome-wide specificity of dCpf1 cytidine base
editors. Nat Commun 11(1):4072. https:/
/doi.
org/10.1038/s41467-020-17889-9
At last, duringan expedition to Vesuvius, Oswald speaks. He had
been at one time in love with an unworthy Frenchwoman; had
lingered in France when his father required his presence in England,
and had finally returned, only to find him dead. From that hour he
had known no peace; remorse had pursued him; his filial love, which
was morbidly excessive, caused him to look upon himself as almost
a parricide, and he considered that he was thenceforward morally
bound to do nothing which his father might disapprove. This absurd
conclusion afflicts Corinne visibly, and the sight of her agitation
reawakens all Oswald’s doubts. He conjures her to tell him her
history. She consents; but begs for a few days’ grace, and employs
the interval in planning and carrying out a fête on Cape Misenum. In
front of the azure, tideless sea she takes her lyre and pours out an
improvisation on the past glories of that classic shore. This, although
Oswald does not know it, is an adieu to her past life, for she
foresees that what she has to tell him of herself will entirely change
her destiny. Either he will refuse to marry her, and then she will
never know happiness again, but wingless, voiceless, will go down to
her tomb, or else he will make her his wife, and the Sibyl will be lost
in the peeress.
The next day she leaves with him the narrative of her youth. She
is the daughter of Lord Edgermond by an Italian wife, consequently
the half-sister of Lucile. At the age of fifteen she had gone to
England, and fallen under the rule of her stepmother, Lady
Edgermond, a cold and rigid Englishwoman, who cared for nothing
outside her small provincial town, and regarded genius as a
dangerous eccentricity. In the narrow monotony of the life imposed
upon her Corinne nearly died. At the age of twenty-one she finally
escaped and returned to Italy, having dropped her family name out
of respect for Lady Edgermond’s feelings. Until her meeting with
Oswald she had led the life of a muse, singing, dancing, playing,
improvising for the whole of Roman society to admire, and had
conceived no idea of greater felicity until learning to love. This love
had been a source of peculiar torment to her from the fact of her
divining how much the unconventionality of her conduct, when fully
57.
known to him,must shock Oswald’s English notions of propriety. In
the first moment, however, his love triumphs over these
considerations, and he resolves to marry Corinne. Only he wishes
first—in order that no reproach may attach to her—to force Lady
Edgermond once again to acknowledge her as her husband’s
daughter. He goes to England, partly for this purpose, partly because
his regiment has been ordered on active service.
In England he again meets Lucile, a cold-mannered, correct, pure-
minded, but secretly ardent English girl, with an odd resemblance in
many ways to a French jeune fille. He mentions the subject of her
step-daughter to the upright but selfish Lady Edgermond, who has
set her heart on seeing Oswald the husband of Lucile. She is too
honorable to try and detach him from Corinne by any underhand
means, but does what she knows will be far more effectual; that is,
she makes him acquainted with the fact that his father had seen
Corinne in her early girlhood, had admired her, but had strongly
pronounced against the marriage proposed by Lord Edgermond
between her and Oswald. In the view of the late Lord Nelvil, she was
too brilliant and distinguished for domestic life. This is a terrible blow
to Oswald. He begins to think he must give up Corinne, and is
strengthened in the idea by perceiving that the beautiful and
virtuous Lucile is in love with him. Finally he marries her, decided at
the last by Corinne’s inexplicable silence. She has not answered his
letters for a month, and he concludes that she has forgotten him.
But her silence is owing to her having left Venice and come to
England. She loses a whole month in London, for very insufficient
reasons—necessary, however, to the story—and at last follows
Oswald to Scotland just in time to learn that he is married, to fall
senseless on the road-side, and to be picked up by the Count
D’Erfeuil. She returns heart-broken to Italy, and dies slowly through
four long years of unbroken misery.
When she is near her end Oswald comes to Florence,
accompanied by his wife and child. He had begun to regret Corinne
as soon as he had married Lucile, who, on her side, being naturally
resentful, takes refuge in coldness and reserve. As soon as Lord
58.
Nelvil learns thathis old love is in Florence and dying he wishes
ardently to see her, but she refuses to receive him. He sends the
child to her, and she teaches it some of her accomplishments. Lucile
visits her secretly, and is converted by her eloquence to the
necessity of rendering herself more attractive to her husband by
displaying some graces of mind.
At last Corinne consents to see Oswald once again, but it shall be,
she determines, in public. This is one of the most unnatural scenes
in the book. Corinne invites all her friends to assemble in a lecture
hall. Thither she has herself transported and placed in an arm-chair.
A young girl clad in white and crowned with flowers recites the Song
of the Swan, or adieu to life, which Corinne has composed, while
Oswald, listening to it and gazing on the dying poetess from his
place in the crowd, is suffocated with emotion and finally faints. A
few days later Corinne dies, her last act being to point with her
diaphanous hand to the moon, which is partially obscured by a band
of cloud such as she and Lord Nelvil had once seen when in Naples.
Even as a picture of Italy, Corinne leaves much to be desired.
Madame de Staël’s ideas of art were acquired. She had no
spontaneous admiration even for the things she most warmly
praised, and her judgments were conventional and essentially cold.
Some of the descriptions are good in the sense of being accurate
and forcibly expressed. But even in the best of them—that of
Vesuvius—one feels the effort. Madame de Staël is wide-eyed and
conscientious, but has no flashes of inspired vision. She can
catalogue but not paint. A certain difficulty in saying enough on
æsthetic subjects is rendered evident by her vice of moralizing.
Instead of admiring a marble column as a column, or a picture as a
picture, she finds in it food for reflection on the nature of man and
the destiny of the world. Some of her remarks on Italian character
are extremely clever, and show her usual surprising power of
observation; but they are generally superficial.
This was due, in part, to her system of explaining everything by
race and political institutions, in part to her passion for
59.
generalization. Because Italianshad produced the finest art and
some of the finest music; because they had no salons and wrote
sonnets; because they had developed a curiously systematic form of
conjugal infidelity; finally, because they had no political liberty,
Madame de Staël constructed a theory which represented them as
simply passionate, romantic, imaginative and indulgent. This theory
has cropped up now and again in literature from her days to our
own, and if partially correct, overlooks the subtler shades and
complex contradictions of the Italian mind.
Roman society in the beginning of this century was far from being
the transfigured and exotic thing represented in Corinne. The
modern Sibyl’s prototype, poor Maddalena Maria Morelli, was
mercilessly pasquinaded, and on her road to the Capitol pelted with
rotten eggs. This gives a very good idea of the sort of impression
that would have been produced on a real Prince of Castel-Forte and
his fellows by the presence in their midst of a young and beautiful
woman, unmarried, nameless, and rich. Corinne’s lavish exhibition of
her accomplishments is another “false note,” as singing and dancing
were but rarely, if ever, performed by amateurs in Italy. What
redeems the book are the detached sentences of thought that gem
almost every page of it. Madame de Staël had gradually shaken off
the vices of style which her warmest admirers deplore in her, and in
her Allemagne she was presently to reveal herself as singularly lucid,
brilliant, and acute. This work of hers on Germany is, perhaps, the
most satisfactory of her many productions. As a review of society,
art, literature, and philosophy, it naturally lends itself to the form
best suited to her essentially analytical mind.
Madame de Staël was always obliged to generalize, that being a
law of her intelligence, and this disposition is accentuated in the
Allemagne, through her desire to establish such contrasts between
Germany and France, as would inspire the latter with a sense of its
defects. She saw Germany on the eve of a great awakening, and
was not perhaps as fully conscious of this as she might have been.
As Saint Beuve happily says, she was not a poet, and it is only poets
who, like birds of passage, feel a coming change of season.
60.
Germany appealed toher, however, through everything in herself
that was least French; her earnestness, her vague but ardent
religious tendencies, her spiritualism, her excessive admiration of
intellectual pursuits. She was, therefore, exceptionally well-qualified
to reveal to her own countrymen the hitherto unknown or
unappreciated beauties of the German mind.
She was, on the other hand, extremely alive to the dullness of
German, and especially of Viennese, society, and portrays it in a
series of delightfully witty phrases. The Allemagne is indeed the
wittiest of all her works, and abounds in the happiest touches.
The opinions expressed on German literature are favorable
towards it, and on the whole correct. If she betrays that Schiller was
personally more sympathetic to her than Goethe, she nevertheless
was quick to perceive in the latter the strain of southern passion, the
light, warmth and color, which made his intellect less national than
universal.
Her chapters on Kant and German philosophy generally, are
luminous if not exhaustive. She takes the moral sentiment as her
standpoint, and pronounces from that on the different systems.
Needless to say, she admires metaphysical speculations, and
considers them as valuable in developing intellect and strengthening
character.
Les Dix Années d’Exil is a charming book. Apart from its interest
as a transcript of the writer’s impressions during her exile at Coppet
and subsequent flight across Europe, it contains brilliant pictures of
different lands, and especially Russia. One is really amazed to note
how much she grasped of the national characteristics during her
brief sojourn in that country. The worst reproach that can be
addressed to her description is that, as usual, it is rather too
favorable. Her anxiety to prove that no country could flourish, during
a reign such as Napoleon’s, made her disposed to see through rose-
colored spectacles the Governments which found force to resist him.
61.
The Considerations onthe French Revolution were published
posthumously. According to Sainte Beuve, this is the finest of
Madame de Staël’s works. “Her star,” he says, “rose in its full
splendor only above her tomb.” It is difficult to pronounce any
summary judgment on this book, which is partly biographical and
partly historical. The first volume is principally devoted to a
vindication of Necker; the second to an attack on Napoleon; the
third to a study of the English Constitution and the applicability of its
principles to France. The first two volumes alone were revised by the
authoress before her death. We find in this work all Madame de
Staël’s natural and surprising power of comprehension. She handles
difficult political problems with an ease that would be more
astonishing still, had the book more unity. As it is, each separate
circumstance is related and explained admirably, but one is not
made to reach the core of the stupendous event of which Europe
still feels the vibration. Her portrait of Napoleon is unsurpassable for
force and irony, for sarcasm and truth. All she possessed of
epigrammatic power seems to have come unsought to enable her to
avenge herself on the mean, great man who had feared her enough
to exile and persecute her.
In closing this rapid review of her works, one asks why was
Madame de Staël not a greater writer? The answer is easy; she
lacked high creative power and the sense of form. Her mind was
strong of grasp and wide in range, but continuous effort fatigued it.
She could strike out isolated sentences alternately brilliant,
exhaustive and profound, but she could not link them to other
sentences so as to form an organic page. Her thought was definite
singly, but vague as a whole. She always saw things separately, and
tried to unite them arbitrarily, and it is generally difficult to follow out
any idea of hers from its origin to its end. Her thoughts are like
pearls of price profusely scattered, or carelessly strung together, but
not set in any design. On closing one of her books, the reader is left
with no continuous impression. He has been dazzled and delighted,
enlightened also by flashes; but the horizons disclosed have
vanished again, and the outlook is enriched by no new vistas.
62.
Then she wasdeficient in the higher qualities of imagination. She
could analyze but not characterize; construct but not create. She
could take one defect like selfishness, or one passion like love, and
display its workings; or she could describe a whole character, like
Napoleon’s, with marvellous penetration; but she could not make her
personages talk or act like human beings. She lacked pathos, and
had no sense of humor. In short, hers was a mind endowed with
enormous powers of comprehension, and an amazing richness of
ideas, but deficient in perception of beauty, in poetry, and true
originality. She was a great social personage, but her influence on
literature was not destined to be lasting, because, in spite of
foreseeing much, she had not the true prophetic sense of
proportion, and confused the things of the present with those of the
future—the accidental with the enduring.
THE END.
63.
Messrs. Roberts Brothers’Publications.
FAMOUS WOMEN SERIES.
GEORGE SAND.
By BERTHA THOMAS.
One volume. 16mo. Cloth. Price, $1.00.
“Miss Thomas has accomplished a difficult task with as much good sense as
good feeling. She presents the main facts of George Sand’s life, extenuating
nothing, and setting naught down in malice, but wisely leaving her readers to
form their own conclusions. Everybody knows that it was not such a life as the
women of England and America are accustomed to live, and as the worst of
men are glad to have them live.… Whatever may be said against it, its result on
George Sand was not what it would have been upon an English or American
woman of genius.”—New York Mail and Express.
“This is a volume of the ‘Famous Women Series,’ which was begun so well
with George Eliot and Emily Brontë. The book is a review and critical analysis of
George Sand’s life and work, by no means a detailed biography. Amantine
Lucile Aurore Dupin, the maiden, or Mme. Dudevant, the married woman, is
forgotten in the renown of the pseudonym George Sand.
“Altogether, George Sand, with all her excesses and defects, is a
representative woman, one of the names of the nineteenth century. She was
great among the greatest, the friend and compeer of the finest intellects, and
Miss Thomas’s essay will be a useful and agreeable introduction to a more
extended study of her life and works.”—Knickerbocker.
“The biography of this famous woman, by Miss Thomas, is the only one in
existence. Those who have awaited it with pleasurable anticipation, but with
some trepidation as to the treatment of the erratic side of her character, cannot
fail to be pleased with the skill by which it is done. It is the best production on
George Sand that has yet been published. The author modestly refers to it as a
sketch, which it undoubtedly is, but a sketch that gives a just and
64.
discriminating analysis ofGeorge Sand’s life, tastes, occupations, and of the
motives and impulses which prompted her unconventional actions, that were
misunderstood by a narrow public. The difficulties encountered by the writer in
describing this remarkable character are shown in the first line of the opening
chapter, which says, ‘In naming George Sand we name something more
exceptional than even a great genius.’ That tells the whole story.
Misconstruction, condemnation, and isolation are the penalties enforced upon
the great leaders in the realm of advanced thought, by the bigoted people of
their time. The thinkers soar beyond the common herd, whose soul-wings are
not strong enough to fly aloft to clearer atmospheres, and consequently they
censure or ridicule what they are powerless to reach. George Sand, even to a
greater extent than her contemporary, George Eliot, was a victim to ignorant
social prejudices, but even the conservative world was forced to recognize the
matchless genius of these two extraordinary women, each widely different in
her character and method of thought and writing.… She has told much that is
good which has been untold, and just what will interest the reader, and no
more, in the same easy, entertaining style that characterizes all of these
unpretentious biographies.”—Hartford Times.
Sold everywhere. Mailed, post-paid, on receipt of price, by the publishers,
ROBERTS BROTHERS, Boston.
65.
Messrs. Roberts Brothers’Publications.
FAMOUS WOMEN SERIES.
MARY LAMB.
By ANNE GILCHRIST.
One volume. 16mo. Cloth. Price, $1.00.
“The story of Mary Lamb has long been familiar to the readers of Elia, but
never in its entirety as in the monograph which Mrs. Anne Gilchrist has just
contributed to the Famous Women Series. Darkly hinted at by Talfourd in his
Final Memorials of Charles Lamb, it became better known as the years went on
and that imperfect work was followed by fuller and franker biographies,—
became so well known, in fact, that no one could recall the memory of Lamb
without recalling at the same time the memory of his sister.”—New York Mail
and Express.
“A biography of Mary Lamb must inevitably be also, almost more, a biography
of Charles Lamb, so completely was the life of the sister encompassed by that
of her brother; and it must be allowed that Mrs. Anne Gilchrist has performed a
difficult biographical task with taste and ability.… The reader is at least likely to
lay down the book with the feeling that if Mary Lamb is not famous she
certainly deserves to be, and that a debt of gratitude is due Mrs. Gilchrist for
this well-considered record of her life.”—Boston Courier.
“Mary Lamb, who was the embodiment of everything that is tenderest in
woman, combined with this a heroism which bore her on for a while through
the terrors of insanity. Think of a highly intellectual woman struggling year after
year with madness, triumphant over it for a season, and then at last
succumbing to it. The saddest lines that ever were written are those descriptive
of this brother and sister just before Mary, on some return of insanity, was to
leave Charles Lamb. ‘On one occasion Mr. Charles Lloyd met them slowly pacing
together a little foot-path in Hoxton Fields, both weeping bitterly, and found, on
66.
joining them, thatthey were taking their solemn way to the accustomed
asylum.’ What pathos is there not here?”—New York Times.
“This life was worth writing, for all records of weakness conquered, of pain
patiently borne, of success won from difficulty, of cheerfulness in sorrow and
affliction, make the world better. Mrs. Gilchrist’s biography is unaffected and
simple. She has told the sweet and melancholy story with judicious sympathy,
showing always the light shining through darkness.”—Philadelphia Press.
Sold by all Booksellers. Mailed, post-paid, on receipt of the
price, by the Publishers,
ROBERTS BROTHERS, Boston.
67.
Messrs. Roberts Brothers’Publications.
Famous Women Series.
RACHEL.
By Mrs. NINA H. KENNARD.
One Volume. 16mo. Cloth. Price, $1.00.
“Rachel, by Nina H. Kennard, is an interesting sketch of the famous woman
whose passion and genius won for her an almost unrivalled fame as an actress.
The story of Rachel’s career is of the most brilliant success in art and of the
most pathetic failure in character. Her faults, many and grievous, are
overlooked in this volume, and the better aspects of her nature and history are
recorded.”—Hartford Courant.
“The book is well planned, has been carefully constructed, and is pleasantly
written.”—The Critic.
“The life of Mlle. Élisa Rachel Félix has never been adequately told, and the
appearance of her biography in the ‘Famous Women Series’ of Messrs. Roberts
Brothers will be welcomed.… Yet we must be glad the book is written, and
welcome it to a place among the minor biographies; and because there is
nothing else so good, the volume is indispensable to library and study.”—Boston
Evening Traveller.
“Another life of the great actress Rachel has been written. It forms part of
the ‘Famous Women Series,’ which that firm is now bringing out, and which
already includes eleven volumes. Mrs. Kennard deals with her subject much
more amiably than one or two of the other biographers have done. She has
none of those vindictive feelings which are so obvious in Madame B.’s narrative
of the great tragedienne. On the contrary, she wants to be fair, and she
probably is as fair as the materials which came into her possession enabled her
to be. The endeavor has been made to show us Rachel as she really was, by
relying to a great extent upon her letters.… A good many stories that we are
68.
familiar with arerepeated, and some are contradicted. From first to last,
however, the sympathy of the author is ardent, whether she recounts the
misery of Rachel’s childhood, or the splendid altitude to which she climbed
when her name echoed through the world and the great ones of the earth vied
in doing her homage. On this account Mrs. Kennard’s book is a welcome
addition to the pre-existing biographies of one of the greatest actresses the
world ever saw.”—N.Y. Evening Telegram.
Sold everywhere. Mailed postpaid, by the Publishers,
ROBERTS BROTHERS, Boston.
69.
Messrs. Roberts Brothers’Publications.
FAMOUS WOMEN SERIES.
THE COUNTESS OF ALBANY.
BY VERNON LEE.
One volume. 16mo. Cloth. Price $1.00.
“It is no disparagement to the many excellent previous sketches to say that
‘The Countess of Albany,’ by Vernon Lee, is decidedly the cleverest of the series
of biographies of ‘Famous Women,’ published in this country by Roberts
Brothers, Boston. In the present instance there is a freer subject, a little farther
removed from contemporary events, and sufficiently out of the way of prejudice
to admit of a lucid handling. Moreover, there is a trained hand at the work, and
a mind not only familiar with and in sympathy with the character under
discussion, but also at home with the ruling forces of the eighteenth century,
which were the forces that made the Countess of Albany what she was. The
biography is really dual, tracing the life of Alfieri, for twenty-five years the heart
and soul companion of the Countess, quite as carefully as it traces that of the
fixed subject of the sketch.”—Philadelphia Times.
“To be unable altogether to acquiesce in Vernon Lee’s portrait of Louise of
Stolberg does not militate against our sense of the excellence of her work. Her
pictures of eighteenth-century Italy are definite and brilliant. They are instinct
with a quality that is akin to magic.”—London Academy.
“In the records of famous women preserved in the interesting series which
has been devoted to such noble characters as Margaret Fuller, Elizabeth Fry,
and George Eliot, the life of the Countess of Albany holds a unique place.
Louise of Albany, or Louise R., as she liked to sign herself, possessed a
character famed, not for domestic virtues, nor even for peculiar wisdom and
creative power, but rather notorious for an easy-going indifference to
conventionality and a worldly wisdom and cynicism. Her life, which is a singular
exponent of the false ideas prevalent upon the subject of love and marriage in
70.
the eighteenth century,is told by Vernon Lee in a vivid and discriminating
manner. The biography is one of the most fascinating, if the most sorrowful, of
the series.”—Boston Journal.
“She is the first really historical character who has appeared on the literary
horizon of this particular series, her predecessors having been limited to purely
literary women. This brilliant little biography is strongly written. Unlike
preceding writers—German, French, and English—on the same subject, the
author does not hastily pass over the details of the Platonic relations that
existed between the Countess and the celebrated Italian poet ‘Alfieri.’ In this
biography the details of that passionate friendship are given with a fidelity to
truth, and a knowledge of its nature, that is based upon the strictest and most
conscientious investigation, and access to means heretofore unattainable to
other biographers. The history of this friendship is not only exceedingly
interesting, but it presents a fascinating psychological study to those who are
interested in the metaphysical aspect of human nature. The book is almost as
much of a biography of ‘Alfieri’ as it is of the wife of the Pretender, who
expected to become the Queen of England.”—Hartford Times.
Sold by all booksellers. Mailed, postpaid, on receipt of the price,
by the publishers,
ROBERTS BROTHERS, Boston.
71.
MESSRS. ROBERTS BROTHERS’
PUBLICATIONS.
FamousWomen Series.
GEORGE ELIOT.
By MATHILDE BLIND.
One vol. 16mo. Cloth. Price, $1.00.
“Messrs. Roberts Brothers begin a series of Biographies of Famous Women
with a life of George Eliot, by Mathilde Blind. The idea of the series is an
excellent one, and the reputation of its publishers is a guarantee for its
adequate execution. This book contains about three hundred pages in open
type, and not only collects and condenses the main facts that are known in
regard to the history of George Eliot, but supplies other material from personal
research. It is agreeably written, and with a good idea of proportion in a
memoir of its size. The critical study of its subject’s works, which is made in the
order of their appearance, is particularly well done. In fact, good taste and
good judgment pervade the memoir throughout.”—Saturday Evening Gazette.
“Miss Blind’s little book is written with admirable good taste and judgment,
and with notable self-restraint. It does not weary the reader with critical
discursiveness, nor with attempts to search out high-flown meanings and
recondite oracles in the plain ‘yea’ and ‘nay’ of life. It is a graceful and
unpretentious little biography, and tells all that need be told concerning one of
the greatest writers of the time. It is a deeply interesting if not fascinating
woman whom Miss Blind presents,” says the New York Tribune.
“Miss Blind’s little biographical study of George Eliot is written with sympathy
and good taste, and is very welcome. It gives us a graphic if not elaborate
sketch of the personality and development of the great novelist, is particularly
full and authentic concerning her earlier years, tells enough of the leading
motives in her work to give the general reader a lucid idea of the true drift and
72.
purpose of herart, and analyzes carefully her various writings, with no attempt
at profound criticism or fine writing, but with appreciation, insight, and a clear
grasp of those underlying psychological principles which are so closely
interwoven in every production that came from her pen.”—Traveller.
“The lives of few great writers have attracted more curiosity and speculation
than that of George Eliot. Had she only lived earlier in the century she might
easily have become the centre of a mythos. As it is, many of the anecdotes
commonly repeated about her are made up largely of fable. It is, therefore,
well, before it is too late, to reduce the true story of her career to the lowest
terms, and this service has been well done by the author of the present
volume.”—Philadelphia Press.
Sold by all booksellers, or mailed, post-paid, on receipt of price,
by the publishers,
ROBERTS BROTHERS, Boston.
73.
Messrs. Roberts Brothers’Publications.
FAMOUS WOMEN SERIES.
EMILY BRONTË.
By A. MARY F. ROBINSON.
One vol. 16mo. Cloth. Price, $1.00.
“Miss Robinson has written a fascinating biography.… Emily Brontë is
interesting, not because she wrote ‘Wuthering Heights,’ but because of her
brave, baffled, human life, so lonely, so full of pain, but with a great hope
shining beyond all the darkness, and a passionate defiance in bearing more
than the burdens that were laid upon her. The story of the three sisters is
infinitely sad, but it is the ennobling sadness that belongs to large natures
cramped and striving for freedom to heroic, almost desperate, work, with little
or no result. The author of this intensely interesting, sympathetic, and eloquent
biography, is a young lady and a poet, to whom a place is given in a recent
anthology of living English poets, which is supposed to contain only the best
poems of the best writers.”—Boston Daily Advertiser.
“Miss Robinson had many excellent qualifications for the task she has
performed in this little volume, among which may be named, an enthusiastic
interest in her subject and a real sympathy with Emily Brontë’s sad and heroic
life. ‘To represent her as she was,’ says Miss Robinson, ‘would be her noblest
and most fitting monument.’ … Emily Brontë here becomes well known to us
and, in one sense, this should be praise enough for any biography.”—New York
Times.
“The biographer who finds such material before him as the lives and
characters of the Brontë family need have no anxiety as to the interest of his
work. Characters not only strong but so uniquely strong, genius so supreme,
misfortunes so overwhelming, set in its scenery so forlornly picturesque, could
not fail to attract all readers, if told even in the most prosaic language. When
we add to this, that Miss Robinson has told their story not in prosaic language,
74.
but with aliterary style exhibiting all the qualities essential to good biography,
our readers will understand that this life of Emily Brontë is not only as
interesting as a novel, but a great deal more interesting than most novels. As it
presents most vividly a general picture of the family, there seems hardly a
reason for giving it Emily’s name alone, except perhaps for the masterly
chapters on ‘Wuthering Heights,’ which the reader will find a grateful
condensation of the best in that powerful but somewhat forbidding story. We
know of no point in the Brontë history—their genius, their surroundings, their
faults, their happiness, their misery, their love and friendships, their
peculiarities, their power, their gentleness, their patience, their pride,—which
Miss Robinson has not touched upon with conscientiousness and sympathy.”—
The Critic.
“‘Emily Brontë’ is the second of the ‘Famous Women Series,’ which Roberts
Brothers, Boston, propose to publish, and of which ‘George Eliot’ was the initial
volume. Not the least remarkable of a very remarkable family, the personage
whose life is here written, possesses a peculiar interest to all who are at all
familiar with the sad and singular history of herself and her sister Charlotte.
That the author, Miss A. Mary F. Robinson, has done her work with minute
fidelity to facts as well as affectionate devotion to the subject of her sketch, is
plainly to be seen all through the book.”—Washington Post.
Sold by all Booksellers, or mailed, post-paid, on receipt of price,
by the Publishers,
ROBERTS BROTHERS, Boston.
75.
Messrs. Roberts Brothers’Publications.
Famous Women Series.
MARGARET FULLER.
By JULIA WARD HOWE.
One volume. 16mo. Cloth. Price $1.00.
“A memoir of the woman who first in New England took a position of moral
and intellectual leadership, by the woman who wrote the Battle Hymn of the
Republic, is a literary event of no common or transient interest. The Famous
Women Series will have no worthier subject and no more illustrious biographer.
Nor will the reader be disappointed,—for the narrative is deeply interesting and
full of inspiration.”—Woman’s Journal.
“Mrs. Julia Ward Howe’s biography of Margaret Fuller, in the Famous Women
Series of Messrs. Roberts Brothers, is a work which has been looked for with
curiosity. It will not disappoint expectation. She has made a brilliant and an
interesting book. Her study of Margaret Fuller’s character is thoroughly
sympathetic; her relation of her life is done in a graphic and at times a
fascinating manner. It is the case of one woman of strong individuality depicting
the points which made another one of the most marked characters of her day.
It is always agreeable to follow Mrs. Howe in this; for while we see marks of
her own mind constantly, there is no inartistic protrusion of her personality. The
book is always readable, and the relation of the death-scene is thrillingly
impressive.”—Saturday Gazette.
“Mrs. Julia Ward Howe has retold the story of Margaret Fuller’s life and career
in a very interesting manner. This remarkable woman was happy in having
James Freeman Clarke, Ralph Waldo Emerson, and William Henry Channing, all
of whom had been intimate with her and had felt the spell of her extraordinary
personal influence, for her biographers. It is needless to say, of course, that
76.
Welcome to ourwebsite – the perfect destination for book lovers and
knowledge seekers. We believe that every book holds a new world,
offering opportunities for learning, discovery, and personal growth.
That’s why we are dedicated to bringing you a diverse collection of
books, ranging from classic literature and specialized publications to
self-development guides and children's books.
More than just a book-buying platform, we strive to be a bridge
connecting you with timeless cultural and intellectual values. With an
elegant, user-friendly interface and a smart search system, you can
quickly find the books that best suit your interests. Additionally,
our special promotions and home delivery services help you save time
and fully enjoy the joy of reading.
Join us on a journey of knowledge exploration, passion nurturing, and
personal growth every day!
ebookbell.com


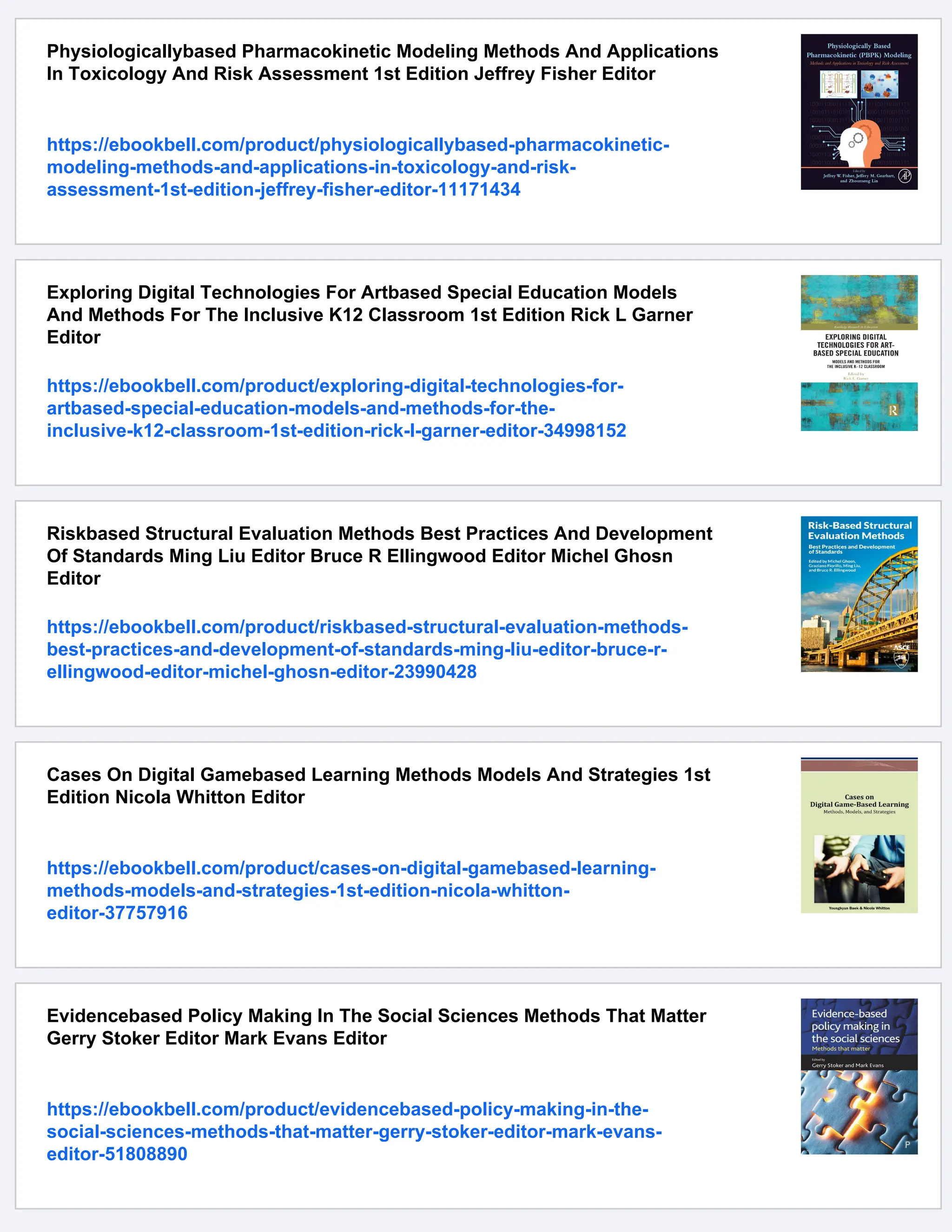














![Chapter 1
Introduction and Perspectives of DNA Base Editors
Beomjong Song and Sangsu Bae
Abstract
DNA base editors, one of the CRISPR-based genome editing tools, can induce targeted point mutations at
desired sites. Their superiority is based on the fact that they can perform efficient and precise gene editing
without generating a DNA double-strand break (DSB) or requiring a donor DNA template. Since they
were first developed, significant efforts have been made to improve DNA base editors in order to overcome
problems such as off-target edits on DNA/RNA and bystander mutations in editing windows. Here, we
provide an overview of DNA base editors with a summary about the history of development of DNA base
editors and report on efforts to improve them.
Key words CRISPR, Base editor, CBE, ABE
1 Introduction
By repurposing the adaptive immune system of bacteria, clustered
regularly interspaced short palindromic repeats (CRISPR), a new
era has opened in the field of genome engineering. The CRISPR
system is superior to other systems such as zinc-finger nucleases
(ZFNs) and transcription activator-like effector nucleases
(TALENs), due to its ability to perform efficient gene targeting in
a programmable, RNA-guided manner. Since it was first proven
that site-specific editing of the human genome can be performed
using the CRISPR system [1–5], diverse genome editing tools
based on the CRISPR system, which enable gene disruption, recov-
ery of the mutated genes, and regulation of the gene expression,
have been introduced.
Nevertheless, there are some fundamental disadvantages asso-
ciated with use of conventional CRISPR-based DNA endonu-
cleases. Gene editing using the CRISPR nucleases initiates with
introduction of DNA double-stranded breaks (DSBs). DSBs are
necessary for both gene insertion and gene disruption, however,
they can also cause unwanted results such as large chromosomal
deletions or genomic rearrangements [6]. In addition, DSB may
Sangsu Bae and Beomjong Song (eds.), Base Editors: Methods and Protocols,
Methods in Molecular Biology, vol. 2606, https://doi.org/10.1007/978-1-0716-2879-9_1,
© The Author(s), under exclusive license to Springer Science+Business Media, LLC, part of Springer Nature 2023
3](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-18-2048.jpg)
![4 Beomjong Song and Sangsu Bae
interrupt precise gene editing by inducing the p53-mediated DNA
damage response [7, 8]. Repair of DSB is a key step in gene editing
using conventional CRISPR nucleases, and there are three path-
ways of DSB repair: homology-directed repair (HDR),
non-homologous end joining (NHEJ), and microhomology-
mediated end joining [9–12]. With HDR, precise DNA edits can
be installed by using a donor DNA template, however it is highly
confined to a specific cell cycle phase, S and G2 phases. On the
other hand, operation of the NHEJ and MMEJ pathways is inde-
pendent of cell cycle phases. However, their application is limited
due to error-prone editing results [6, 13].
Alternatively, DNA base editors can edit a gene at a single
nucleotide level with accompanying little DSB generation
[14, 15]. In addition, DNA base editors can be used in both
dividing and non-dividing cells because their operation is not lim-
ited to a specific cell cycle phase. The fact that use of donor
templates is unnecessary also makes DNA base editors even more
of a standout.
2 The Construction of Cytosine Base Editors (CBEs) and the Mechanism of Cytosine
Base Editing
Cytosine base editors (CBEs) commonly contain a fusion form of a
cytidine deaminase and a Cas effector. The initial CBE employed
rAPOBEC1 as a cytidine deaminase and deactivated/dead Cas9
(dCas9) to avoid generation of DSBs (BE1). Recruitment of cyti-
dine deaminase to a target DNA site can occur through an interac-
tion between the dCas9 and the associated single-guide RNA
(sgRNA), with recognition of the single-stranded DNA (ssDNA)
in the R-loop structure formed on the target site. Cytidine deami-
nase then converts deoxycytidines into deoxyuridines. Deoxyuri-
dine preferentially pairs with deoxythymidine, thus the resulting
mismatch U/G pair induces activation of DNA repair pathways
such as base excision repair (BER) and mismatch repair (MMR)
[16]. While the U/G pair can be converted into a T/A pair as
desired through the DNA repair pathways, the U/G pair can be
returned to a C/G pair or non-T/A outcomes can be generated by
the activity of a base excision repair enzyme, uracil N-glycosylase
(UNG) [17]. To reduce such unwanted conversions, uracil glyco-
sylase inhibitor (UGI) peptide was fused to dCas9 (BE2) [14]. In
addition, adoption of Cas9 nickase (nCas9) instead of dCas9
resulted in improved efficiency of base editing by causing the repair
mechanisms to use the strand with deoxyuridine as a template for
repair rather than the opposite strand with a nick (BE3) [14]. For
further suppression of the unwanted conversion, other cytidine
deaminases such as AID (AID-BE3) [18] and PmCDA1 (CDA1-
BE3) [19] were employed instead of rAPOBEC1, or a second UGI](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-19-2048.jpg)
![Introduction and Perspectives of DNA Base Editors 5
was added (BE4) [17]. Improved editing efficiency of BE4 was
achieved through codon optimization (BE4-max) [20] and ances-
tral reconstruction (AncBE4max) [21].
3 The Construction of Adenine Base Editors (ABEs) and the Mechanism of Adenine
Base Editing
The structure and the base editing mechanism of ABEs are similar
to those of CBEs. In ABEs, nCas9 is fused to adenosine deaminase.
All adenosine deaminases discovered so far show activity on RNA
not DNA, therefore, the tRNA-specific adenosine deaminase,
TadA, from Escherichia coli (E. coli) was engineered to operate on
DNA (eTadA*) [15]. ABEs are recruited to the target site of DNA
in the same way as CBEs and convert an adenosine into an inosine,
generating an I/T pair. The mismatched I/T pair is then converted
into a G/C pair through the I/C pair.
The primitive ABE (ABE1.2) comprises the eTadA* monomer.
Because TadAs form a homodimer in nature controlling adenosine
deamination and target binding, respectively, a wild-type TadA
(wtTadA) was fused to eTadA* in the follow-up version of ABE
(ABE2.9). Subsequently, several mutations were introduced into
eTadA* in order to obtain higher editing efficiency in ABE7.9 and
ABE7.10 [15]. Further improvement of ABE7.10 was achieved
through addition of modified NLS and codon optimization in
ABEmax [21]. In the most recent studies, ABE8 variants (ABE8e
and ABE8s) were developed by removing wtTadA and introducing
additional mutations into eTadA* [22, 23].
4 Improvement of DNA Base Editors
4.1 Expanding
Targetable Sites
(Table 1)
Because DNA base editors are based on the CRISPR-Cas system,
targeting DNA sequences is limited by the preference of Cas
nucleases for protospacer adjacent motif (PAM). The original
form of CBEs and ABEs employing Streptococcus pyogenes
(SpCas9) can target only the DNA sequences neighboring the
NGG PAM sequence. Orthologs of Cas effectors having different
PAM preferences were employed, or specific mutations were intro-
duced into the original SpCas9 nucleases in order to expand the
targeting ability of DNA base editors [20, 22–34].
4.2 Modification of
Editing Windows
DNA base editors operate in a confined editing window, which
occasionally results in unwanted base conversion (i.e., bystander
mutation) if multiple Cs or As are located within a editing window.
The frequency of bystander mutations can be reduced by narrowing
the editing window. Several studies have reported that introduction
of mutations in the deaminase determines the width of the editing
window with little effect on the deaminase activity or even with](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-20-2048.jpg)
![Deaminase PAM References
6 Beomjong Song and Sangsu Bae
Table 1
Diverse combinations of deaminases and Cas effectors
Cas
effectors
Cytidine
deaminase
rAPOBEC1 SpCas9 NGG, NG/GAA/GAT, NGA,
NGAG, NGCG, NG
[14, 17, 21, 24, 32,
35, 47–54]
Spy-mac
Cas9
NAA [20]
SaCas9 NNGRRT, NNNRRT [24]
SauriCas9 NNGG [27]
LbCas12a TTTV [28]
AsCas12a TTTV [29]
ScCas9 NNG [31]
hAPBEC3A spCas9 NGG [18, 36, 50, 52]
PmCDA1 spCas9 NGG, NG [18, 19, 26, 37]
cjCas9 NNNVRYAC [55]
hAID spCas9 NGG [18, 56]
PpAPOBEC1 spCas9 NGG [57]
RrA3F SpCas9 NGG [57]
AmAPOBEC1 SpCas9 NGG [57]
SsAPOBEC3B spCas9 NGG [57]
Adenosine
deaminase
TadA7.10 SpCas9 NGG, NGA, NGCG, NG,NG/
GAA/GAT
[15, 21, 25, 30, 32–
34, 47, 48]
Spy-mac
Cas9
NAA [20]
SaCas9 NNGRRT, NNNRRT [32, 33]
ScCas9 NNG [31]
SauriCas9 NNGG [27]
Nme2Cas9 NNNNCC [58]
TadA8e SpCas9 NGG [22, 23]
SaCas9 NNGRRT [22]
LbCas12a TTTV [22]
AsCas12a TTTV [22]
Cas12f TTTR [46]
TadA8s SpCas9 NGG, NG [23]
SaCas9 NNGRRT [23]
TadA8.17 SpCas9 NGG [23]
TadA7.10 SpCas9 NGG [18, 32, 50, 59]
enhanced editing efficiency [14, 24, 28, 35]. Alternatively, deami-
nases requiring a specific motif can be employed as a method of
lowering the number of bystander mutations [36, 37].
On the other hand, expanding the editing window might be
necessary if there is no Cs or As within the editing window. DNA
base editors having circularly permuted SpCas9 (CP-spCas9)
nucleases have shown broaden editing windows without changes
in PAM preference [32]. In addition, some variants of DNA base
editors with CP-SpCas9 can edit bases upstream of the protospacer
on both target and non-target strands [32].](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-21-2048.jpg)
![Introduction and Perspectives of DNA Base Editors 7
4.3 Generating
Transversion Edits
Conventional DNA base editors are limited to DNA base transition
mutations (purine to purine or pyrimidine to pyrimidine), thus
there is a need for novel DNA base editors that can induce transver-
sion mutations. As mentioned above, inhibition of UNG is
required for C-to-T conversion by CBE. In other words, use of
CBE can result in C/G to non-T/A outcomes through BER path-
ways mediated by UNG. Indeed, removal of UGI from CBEs such
as BE4max resulted in an increase of C-to-G edits compared to the
original BE4max. The bias to C-to-G conversion was enhanced by
addition of UNG protein to CBEs [38]. In addition, C-to-A edit-
ing was induced by replacement of rAPOBEC1 with AID in
prokaryotes [39].
4.4 Generating Dual
Base Conversions of
C-to-T and A-to-G at
Once
Efforts to expand the capabilities of base editing led to the devel-
opment of DNA base editors that can introduce both C-to-T and
A-to-G conversions at the same time. According to several studies
reported around the same time, fusion of both adenosine deami-
nase and cytidine deaminase to a single Cas nuclease exhibits a dual
base editing activity with minimal RNA off-target effects [40].
4.5 TC-Specific Base
Editing by ABE
It was revealed that ABE can introduce unwanted C-to-T edits
within the editing window [41]. For its original purpose, a
D108Q mutation was introduced to the adenosine deaminase to
suppress the cytidine deamination activity. Interestingly, P48R
mutation in the adenosine deaminase showed opposite effect;
cytosine following thymine (i.e., TC) was majorly converted. As a
result, the ABE with P48R mutation could induce TC-to-TT and
TC-to-TG editing with and without UGI, respectively
[42]. Because the cytosine editing by the mutated ABE is promi-
nent at the TC motif, this novel DNA base editor can be used as a
TC-specific base editing tool.
4.6 Adeno-
Associated Virus
(AAV)-Loadable BEs
The availability of efficient and safe delivery methods is a prerequi-
site for use of gene editing tools as therapeutics. Among methods
for virus-based delivery, AAV is widely used due to efficient expres-
sion of its cargo with less toxicity such as the integration of viral
DNA into the host genome. Unfortunately, the size of DNA base
editors, which exceeds the packaging capacity of AAV (~4.7 kb),
causes difficulty in loading DNA base editors into a single AAV
vector. In the effort to bypass this problem, one breakthrough was
achieved by splitting DNA base editors into two parts for packaging
into dual AAV vectors [43, 44]. However, the transduction effi-
ciency of the AAV vector is compromised by the dual AAV system
[45]. Alternatively, successful implantation of a small-sized Cas
nuclease such as Cas12f in ABE was achieved, resulting in genera-
tion of dCasMINI-ABE, which can be loaded into a single
AAV [46].](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-22-2048.jpg)




![Chapter 2
Web-Based Computational Tools for Base Editors
Gue-Ho Hwang and Sangsu Bae
Abstract
CRISPR-based base editors are efficient genome editing tools for use in base correction. Currently, there
are various versions and types of base editors with different substitution patterns, editing windows, and
protospacer adjacent motif (PAM) sequences. For the design of target sequences, consideration of
off-target sequences is required. In addition, for assessment of base editing outcomes in bulk populations,
the analysis of high-throughput sequencing data is required. Several web browser-based computation
programs have been developed for the purpose of target design and NGS data analysis, especially for
users with less computational knowledge. In this manuscript, depending on the purpose of each program,
we provide an explanation of useful tools including BE-Designer for design of targets and BE-Analyzer for
analysis of NGS data that were developed by our group, CRISPResso2 for analysis of NGS data developed
by Luca Pinello group, DeepBaseEditor for prediction of target efficiency developed by Hyongbum Henry
Kim group, and BE-Hive for prediction of target outcome developed by David Liu group.
Key words Base editors (BEs), Web tool, Computational tools for base editors
1 Introduction
Base editors are efficient genome editing tools for use in making
nucleotide corrections in specific regions without double-strand
breaks (DSBs) and use of additional donor DNA [1, 2]. CRISPR-
Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats
and CRISPR associated protein 9) [3–5] shows high efficiency for
generation of DSBs in the target sequence [6]; however CRISPR-
Cas9 does not have adequate efficiency for making nucleotide edits
[3–5, 7, 8]. Base editors consist of partially inactive Cas9 (nCas9)
connected with deaminase. By using nCas9, base editors take
advantage of CRISPR-Cas9 system, which has the capacity to effi-
ciently recognize the target sequence and produces single-strand
DNA (ssDNA) by unwinding the DNA strands [9]. Base editors
use deaminase to make a substitution when a single strand of DNA
is exposed. Depending on the type of deaminase, base editors can
classify cytosine base editors (CBEs) [1], which make C-to-T
Sangsu Bae and Beomjong Song (eds.), Base Editors: Methods and Protocols,
Methods in Molecular Biology, vol. 2606, https://doi.org/10.1007/978-1-0716-2879-9_2,
© The Author(s), under exclusive license to Springer Science+Business Media, LLC, part of Springer Nature 2023
13](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-27-2048.jpg)
![14 Gue-Ho Hwang and Sangsu Bae
conversion using cytidine deaminase, and adenine base editors
(ABEs) [2], which make A-to-G conversion using adenosine deam-
inase. C-to-G transversion (C:G to G:C base editors, CGBEs) was
developed in 2021 by Wei Leong Chew’s group using uracil-DNA
glycosylase [10, 11]. Various versions of base editors, such as
BE4max [12], which increases efficiency in mammalian cell types;
CBE-NG [13], which increases coverage of the protospacer adja-
cent motif (PAM) sequence; enAsCas12a-BE [14], which applies
AsCas12a instead of SpCas9; ABE8e [15], which increases the
accuracy of ABEs; and Target-AID [16, 17], which decreases the
range of the editing window, have been developed in an effort to
improve efficiency, accuracy, and window coverage.
Design of target sequences and analysis of NGS data are diffi-
cult without use of computational programs. In addition to the
PAM sequence and termination of the U6 promoter [18], which
are considered when designing the CRISPR-Cas9 target sequence,
the substitution pattern and base editing window according to the
type of base editor are also considered in the design of the target
sequence for base editors. In addition, because Cas9 can recognize
DNA sequences that are similar to the target sequence, off-target
sequences are also considered in the process of design [19, 20].
Therefore, through the adoption of programs for off-target analy-
sis, such as Cas-OFFinder [21] and CROP-IT [22], the program
for use in designing target sequences for base editor, such as
BE-Designer [23], was developed with consideration of off-target
sequences. There are also programs for use in prediction, such as
DeepBaseEditor [24] and BE-Hive [25], which predict the effi-
ciency or outcome of base editors using machine learning prior to
conduct of experiments. In the progress of the analysis, NGS results
should be aligned to a reference sequence and classified according
to their mutation patterns. However, because NGS data include
too many reads, analysis of the data cannot be performed without
use of computational programs. BE-Analyzer [23] and CRIS-
PResso2 [26] were developed for the analysis of NGS results for
base editors.
According to the running environment, there are a variant of
programs that provide only off-line support, that use a web
browser, and that can function under both conditions. Installation
of additional programs or knowledge of the programming language
may be required for use of off-line programs; thus use of off-line
programs is difficult without computational knowledge. Therefore,
the computational programs that use a web browser in design and
analysis of base editor outcome are selectively described in this
manuscript according to the purpose of each program, as follows:
(1) design tool (BE-Designer), (2) efficiency prediction tool
(DeepBaseEditor), (3) outcome prediction tool (BE-Hive), and
(4) analysis tool (BE-analyzer and CRISPResso2).](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-28-2048.jpg)



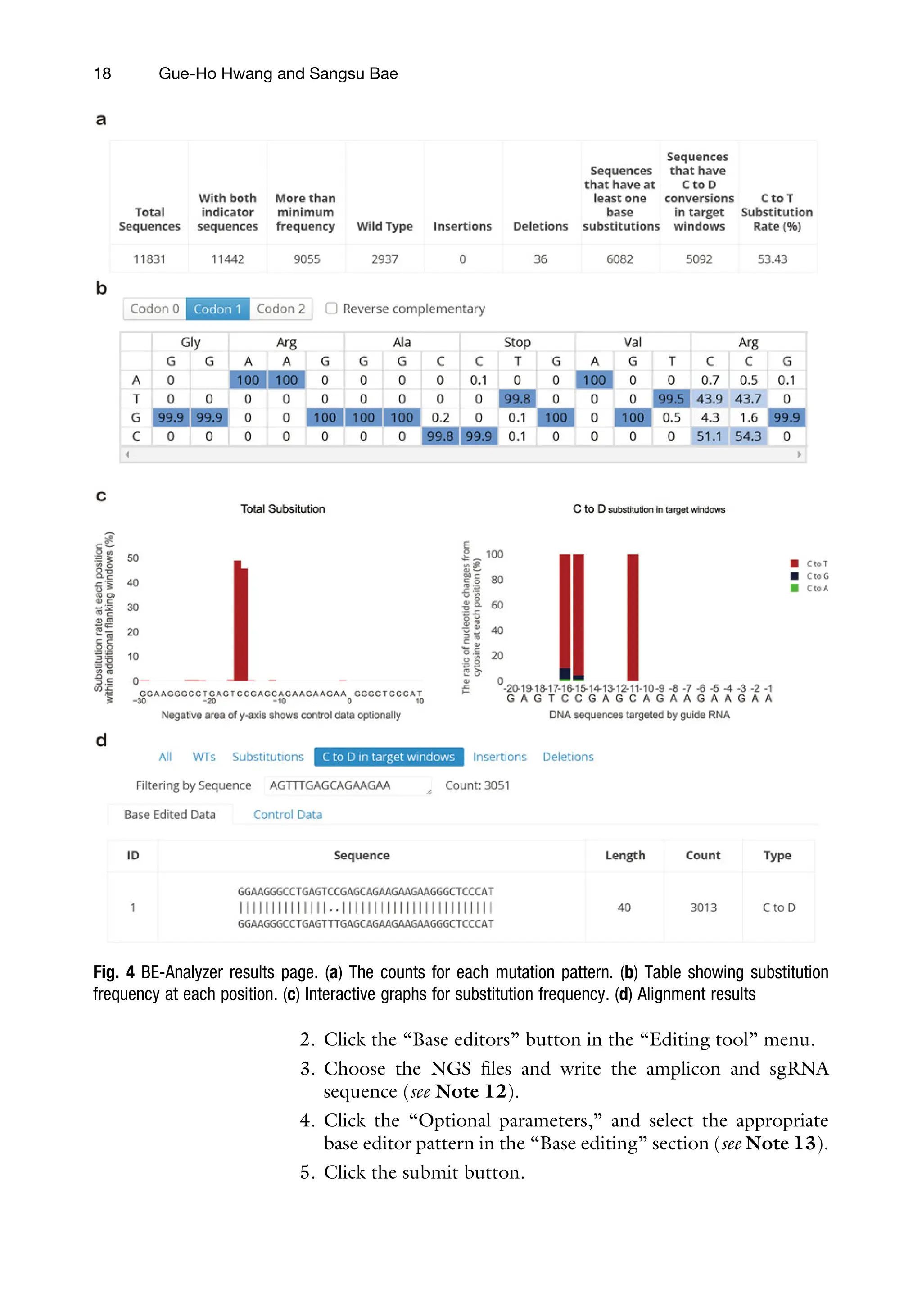



![Chapter 3
Prediction of Base Editing Efficiencies and Outcomes Using
DeepABE and DeepCBE
Jinman Park and Hui Kwon Kim
Abstract
Adenine base editors (ABEs) and cytosine base editors (CBEs) have been widely used to introduce disease-
relevant point mutations at target DNA sites of interest. However, the introduction of point mutations
using base editors can be difficult due to low editing efficiencies and/or the existence of multiple target
nucleotides within the base editing window at the target site. Thus, previous works have relied heavily on
experimentally evaluating the base editing efficiencies and outcomes using time-consuming and labor-
intensive multi-step experimental processes. DeepABE and DeepCBE are deep learning-based computa-
tional models to predict the efficiencies and outcome frequencies of ABE and CBE at given target DNA
sites, in silico. Here, we describe the step-by-step procedure for the accurate determination of specific target
nucleotides for ABE or CBE editing on the online available web tool, (DeepBaseEditor, https:/
/deepcrispr.
info/DeepBaseEditor).
Key words Genome editing. Base editing, Adenine base editor, Cytosine base editor, Deep learning-
based computational model, DeepABE, DeepCBE, DeepBaseEditor
1 Introduction
Adenine base editors (ABEs) [1] and cytosine base editors (CBEs)
[2] can convert A·T base pairs to G·C base pairs and G·C base pairs
to A·T base pairs, respectively, without generating double-stranded
DNA breaks or requiring co-delivery of donor DNA templates.
They enable the efficient modification of disease-relevant mutations
in various species and cell types including human cells. However,
base editor-mediated modification of disease relevant mutations
can be hampered when the base editing efficiency is low and/or
the base editing produces excess of unwanted outcome sequences
especially in cases where multiple target adenines and cytosines are
closely clustered, for ABE and CBE, respectively, within the base-
editable window. Therefore, the examination of editing efficiencies
Sangsu Bae and Beomjong Song (eds.), Base Editors: Methods and Protocols,
Methods in Molecular Biology, vol. 2606, https://doi.org/10.1007/978-1-0716-2879-9_3,
© The Author(s), under exclusive license to Springer Science+Business Media, LLC, part of Springer Nature 2023
23](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-37-2048.jpg)
![24 Jinman Park and Hui Kwon Kim
and editing outcomes of base editing is of utmost importance albeit
through time-consuming experiments including preparation of
single-guide RNA (sgRNA), delivery of the sgRNA-encoding vec-
tor to target cells together with ABE- or CBE-encoding vector, and
PCR amplification of target DNA region followed by deep
sequencing.
Several computational models have been developed for predict-
ing the CRISPR-Cas9 nuclease-directed genome editing efficien-
cies [3–9] or the outcomes [10, 11]. However, these models show
limited performance in predicting base editor activity even at the
same target DNA sequences. DeepABE and DeepCBE are deep
learning-based computational models that accurately predict the
base editing efficiencies and outcomes of ABE and CBE, respec-
tively, without laborious experiments [12]. DeepABE and
DeepCBE predict the base editing efficiencies and outcome fre-
quencies using the features that affect the genome editing efficien-
cies and outcomes at given target sequences. These models are
provided as web tools available on https:/
/deepcrispr.info/
DeepBaseEditor. Here, we have prepared a step-by-step procedure
in procuring the target sequence information as input for the web
tools (Fig. 1) and for interpreting the prediction results (Fig. 2)
which will greatly facilitate genome editing using ABE and CBE.
Fig. 1 The model selection and sequence input page of DeepBaseEditor](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-38-2048.jpg)

![26 Jinman Park and Hui Kwon Kim
and 2) [12]. The wide target sequences should be entered in
FASTA format, starting with a description line followed by
lines of target sequence information. The description line
should begin with “>” symbols to distinguish it from the
following lines with the target sequences. Alternatively, a .zip
file which contains FASTA or FA file can be directly uploaded
to the web server. Examples of input sequence for DeepBaseE-
ditor are as follows:
>example1
ATAACACCGATGACATATACGACAAGTGACGGAGGCA
GATAC
>example2
CTGATTAGACAGATATAAACAGTGGCAAACGTAGG
AAATATAA
3. Submission of the input target sequence: Next, the sequence of
interest can be submitted to the server by clicking the “submit”
button on the web page (Fig. 1c).
4. Find all possible guide RNA sequences within the input sequences:
After the sequence submission, DeepBaseEditor automatically
searches the 5′-NGG-3′ PAM sequences and their
corresponding guide RNA sequences on either DNA strand
and shows the output table containing ID, location, target
sequence, guide RNA sequence, and GC contents of guide
RNA sequences (Fig. 2a). The 5′-NGG-3′ PAM sequence for
each target sequence is shown in red. The ID and location
indicate the description of FASTA format input sequence and
the start:end position of the target sequences within the input
sequence, respectively.
5. Prediction of base editing efficiency for the target sequences:
DeepBaseEditor automatically extracts features including
position-independent and position-dependent nucleotides
and dinucleotides, melting temperature, GC counts, and the
minimum self-folding free energy from the target sequences
(see Note 3). Next, it predicts the base editing efficiencies using
the features and shows the predicted base editing efficiencies on
the output table (Fig. 2a). Here, ABE efficiency and CBE
efficiency refer to the predicted frequency of NGS read counts
(or DNA copies) that contain the intended base-edited
sequences (A to G and C to T conversion for ABE and CBE,
respectively) within the wide editable window (positions 3–10;
see Notes 4 and 5). The output is a downloadable .txt file, and
the user can use the URL shown on the result page for later
access.
6. Prediction of all possible outcome sequences and predicted outcome
frequencies for each target sequences: If there are more than one
target adenine or cytosine in the editable window, base editing](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-40-2048.jpg)
![DeepBaseEditor for Base Editing 27
usually generates diverse outcome sequences. When the user
clicks the ID in each row, DeepBaseEditor provides a table with
all possible outcome sequences, relative proportion (ABE pro-
portion and CBE proportion), and the absolute frequency
(DeepABE and DeepCBE scores) of each outcome sequence
(Fig. 2b; see Notes 6 and 7). The output is a downloadable .txt
file, and the user can use the URL shown on the result page for
later access.
7. Interpretation of prediction scores: DeepABE and DeepCBE
accurately predict the efficiencies and outcome frequencies of
ABE- and CBE-mediated editing at given target sequences
indicated with Pearson’s correlations ranging from 0.50 to
0.95 in HEK293T, HCT116, and U2OS cell lines and
human induced pluripotent stem cells (iPSCs). However, the
absolute editing frequencies of ABE and CBE can vary depend-
ing on the experimental conditions including the cell type,
delivery efficiency, incubation time, and the version of deami-
nase domain (see Notes 8 and 9) [12].
8. Batch run: If there are thousands of target sequences to be
tested, the user may download and use the source codes for
DeepABE and DeepCBE available on Github (https:/
/github.
com/MyungjaeSong/Paired-Library/tree/DeepCRISPR.
info/DeepBaseEditor). The required versions of Python and
various dependent packages are listed below and also available
on the Github page. Briefly, the latest successful run was con-
ducted on Python (2.7.12) using numpy (1.14.5), scipy
(1.1.0), Tensorflow (1.4.1) with CUDA (8.0.61), and
cuDNN (6.0.21). The input target sequences can be listed on
a .txt file using the file format:>example ABE_Efficiency
input1: ./ABE_Efficiency_sample.txt # List of Target
Sequence(s)
Target number 30 bp target sequence (4 bp + 20 bp proto-
spacer + PAM + 3 bp) 1 AACTGAAGGCTGAACAGCA
GGGGTGGGAGA
input2: ./ABE_Efficiency_Weight/ # Pre-trained Weight Files
The input target sequence and pre-trained weight files
should be located in the same working directory as the main
script file, TEST_ABE_Efficiency.py. The user can make modi-
fication to the system paths and parameters by editing the
sections labeled “## System Paths ##” and “## Run Para-
meters ##” within the source code, respectively.
The batch run procedure for CBE is as described above for
ABE with the respective input and script files.>example
CBE_Efficiency
input1: ./CBE_Efficiency_sample.txt # List of Target
Sequence(s)](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-41-2048.jpg)
![28 Jinman Park and Hui Kwon Kim
Target number 30 bp target sequence (4 bp + 20 bp proto-
spacer + PAM + 3 bp) 1 AACTGAAGGCTGAACAGCA
GGGGTGGGAGA
input2: ./CBE_Efficiency_Weight/ # Pre-trained Weight Files
The output file will be generated in a directory named
“output” within the same working directory unless otherwise
modified by the user with the expected file format as below:
>example output
outputs/TEST_OUTPUT_for_test.xlsx
Predicted activity score for sequence 1 : 25.79517365
Following the same principle, detailed outline of the batch run
operation for additional analyses is available on the Github page,
https:/
/github.com/MyungjaeSong/Paired-Library/tree/
DeepCRISPR.info/DeepBaseEditor
4 Notes
1. The optimal length of wide target sequences for DeepABE and
DeepCBE was determined based on the results of cross-
validation. Using 25-nt (2-nt left neighbor + 20-nt guide
sequence binding region + 3-nt PAM sequence) and 24-nt
(1-nt left neighbor + 20-nt guide sequence binding region +
3-nt PAM sequence) wide target sequences showed the highest
performances for the prediction of ABE- and CBE-directed
base editing efficiencies, respectively. The models that use
26-nt (3-nt left neighbor + 20-nt guide sequence binding
region + 3-nt PAM sequence) and 25-nt (2-nt left neighbor +
20-nt guide sequence binding region + 3-nt PAM sequence)
wide target sequences show the highest performances for the
prediction ABE- and CBE-directed base editing outcome pro-
portions, respectively.
2. DeeABE and DeeCBE were developed using data sets of
ABE7.10 (variant of ABE) and BE4 (variant of CBE) mediated
base editing efficiencies and outcome proportions at 13,504
and 14,157 wide target sequences, respectively. ABE7.10 [1]
and BE4 [13] share the Cas9 nickase domain derived from
Streptococcus pyogenes which recognized 5′-NGG-3′ PAM
sequences.
3. Analysis of large-scale data sets allowed for the characterization
of features affecting the ABE and CBE efficiencies and
outcome proportions. These features include position-
independent and position-dependent nucleotides and](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-42-2048.jpg)
![Total reads of a
transitions A T to G
ð Þ
within
R
Total rea
transitions A
ð Þ
w Þ
DeepBaseEditor for Base Editing 29
dinucleotides, melting temperature, GC counts, and the mini-
mum self-folding free energy from the target sequences. Previ-
ous studies have demonstrated that SpCas9 activities were
slightly higher at target sites with high chromatin accessibility
[6]. Comparing DNase I hypersensitive (DHS) sites to
non-DHS sites indicated CBE efficiency was significantly (1.9
fold, P = 0.0077) higher while ABE was less so (1.1 fold,
P = 0.55).
4. We calculated the base editing efficiencies as follows:
Base editing efficiency %
ð Þ =
ll sequences that contain inteanded target nucleotide
C base pairs for ABE : C G to T A base pairs for CBE
the wide editable window at positions 3–10
ð Þ
Total reads
× 100
5. The high-throughput evaluation of base editing efficiencies at
thousands of target sequences showed that the editable win-
dow of ABE (ABE7.10) and CBE (BE4) spans positions 3–10,
although a relatively higher level of base editing can be achieved
at positions 4–8 for both ABE and CBE. Here, position 20 indi-
cates the nucleotide located immediately 5′ upstream of the
PAM sequence.
6. We calculated the base editing outcome proportion as follows:
Base editing outcome proportion =
eads of the specific base‐edited outcome sequence
ds of all sequences that contain intended target nucleotide
T to G C base pairs for ABE; C G to T A base pairs for CBE
ithin the wide editable window at positions 3–10
ð
7. DeepABE and DeepCBE scores indicate the predicted absolute
frequency of each base-edited outcome sequence. Accordingly,
multiplying the base editing efficiency and the base editing
outcome proportion yields the absolute frequency of an out-
come sequence among the total reads (DNA copies). Dee-
pABE and DeepCBE scores are calculated by multiplying the
ABE_proportion and ABE_efficiency or CBE_proportion and
CBE efficiency, respectively.](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-43-2048.jpg)
![Absolute frequency of a
=
Reads of the specific
T
=
Reads
Total reads of all
Transitions A T to
ð Þ
Within the w
×
Total reads of all s
Transitions A T to
ð Þ
Within the w
= base editing outcome
30 Jinman Park and Hui Kwon Kim
n outcome of base‐editing %
ð Þ
base‐edited outcome sequence
otal reads
× %
ð Þ
of the specific base‐edited outcome sequence
sequences that contain intended target nucleotide
G C base pairs for ABE; C G to T A base pairs for CBE
ide editable window at positions 3–10
ð Þ
equences that contain intended target nucleotide
G C base pairs for ABE; C G to T A base pairs for CBE
ide editable window at positions 3–10
ð Þ
Total reads
× 100
proportional × base diting efficiency %
ð Þ
8. Training data sets of DeepABE and DeepCBE were obtained
using the high-throughput evaluation experiments in
HEK293T cells [12]. For the high-throughput evaluation of
base editing activities, a lentiviral library of thousands of guide
RNA-encoding and target sequence pairs that were used previ-
ously for the high-throughput evaluation of Cas9 activities [6]
were adopted. Next, HEK293T cells were transduced with
lentiviral library selected using antibiotics. The cell library was
transiently transfected with either ABE7.10- or BE4-encoding
plasmids. Five days post transfection, cells were harvested and
subjected to deep sequencing to measure the base editing
efficiencies and outcomes at the lentivirally integrated target
sequence. Thus, prediction values of DeepABE and DeepCBE
reflect the absolute editing frequencies of ABE7.10- and
BE4-directed base editing efficiencies measured in HEK293T
cells at 5 days post transfection.
9. Recently, various improved versions of ABE and CBE, involv-
ing biochemical modifications to the deaminase domain, have
been reported [14–20]. However, the Cas9 nickase domain
remains the main catalytic component among these variants.
As such, DeepABE and DeepCBE are expected to accurately
predict the base editing outcomes and efficiencies for these
variants albeit future works can address the potential changes
to the base editing activities elicited by the deaminase domain
modifications.](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-44-2048.jpg)


![Chapter 4
Profiling Genome-Wide Specificity of dCpf1 Cytidine Base
Editors Using Digenome-Seq
Daesik Kim
Abstract
Digenome-seq is a powerful approach for determining the genome-wide specificity of programmable
nuclease including CRISPR-Cas9 and CRISPR-Cpf1 (also known as Cas12a) and programmable deami-
nase including cytosine base editors (CBEs) and adenine base editors (ABEs). To define the genome-wide
specificity of dLbCpf1-BE (also known as dLbCas12a-BE), genomic DNA is first incubated with dLbCpf1-
BE, which induces C-to-U conversion at on-target and off-target sites, and then treated with a mixture of
E. coli uracil DNA glycosylase (UDG) and Endonuclease VIII, which creates single-strand breaks (SSBs) by
removing uracil in vitro. Digested genomic DNA is subjected to WGS, and then sequencing reads are
aligned to the reference genome, resulting in straight alignments at on-target and off-target sites. The
in vitro cleavage sites related to the straight alignments can be identified using the Digenome-seq
computer tool.
Key words Cas12a, Base editor, Genome editing, Off-target, Whole-genome sequencing
1 Introduction
Cytosine base editors (CBEs) and adenine base editors (ABEs) are
CRISPR RNA-guided programmable deaminases made up of a
dCas9 (or nCas9) or dCpf1 and an engineered cytosine or adenine
deaminase [1–5]. Base editors convert C-to-T or A-to-G within the
non-target DNA strand of the target site, without inducing DNA
double-strand breaks (DSBs). Base editors have been widely used
to induce or correct point mutations [6–8].
Catalytically dead Lachnospiraceae bacterium Cpf1 (dLbCpf1,
also known as dLbCas12a)-BE was developed by fusion of
dLbCpf1 with the cytosine deaminase APOBEC1 [4]. dLbCpf1-
BE recognizes TTTV PAM sequences and induces C-to-T conver-
sions at positions 8–13 (numbering in the protospacer from 1 to
23 in the 5′–3′ direction) [4]. However, dLbCpf1-BE can cause
off-target mutations in locations with similar sequences to the
Sangsu Bae and Beomjong Song (eds.), Base Editors: Methods and Protocols,
Methods in Molecular Biology, vol. 2606, https://doi.org/10.1007/978-1-0716-2879-9_4,
© The Author(s), under exclusive license to Springer Science+Business Media, LLC, part of Springer Nature 2023
33](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-47-2048.jpg)
![34 Daesik Kim
target site. To define genome-wide specificity of dLbCpf1-BE
without bias, we developed a modified Digenome-seq, which
could capture single-strand breaks (SSBs) [9]. In this method,
purified genomic DNA (gDNA) is incubated with dLbCpf1-BE
in vitro to induce C-to-U conversion at the on-target and potential
off-target sites, and then uracil is removed using uracil DNA gly-
cosylase (UDG) and DNA glycosylase-lyase endonuclease forming
SSBs in the on-/off-target sites. Digested DNA is fragmented,
end-repaired, and ligated to sequencing adaptors to generate
sequencing libraries. The sequencing libraries are subjected to
whole-genome sequencing (WGS), and sequencing reads are
aligned to a reference genome such as hg19 (Human) and mm10
(Mouse). To profile genome-wide off-target sites of dLbCpf1-BE,
we developed a computer program that could find genome-wide
SSBs without bias [9]. Here, we present a detailed experimental
procedure for profiling the genome-wide specificity of
dLbCpf1-BE.
2 Materials
2.1 crRNA DNA
Templates
1. crRNA_F (Macrogen): 5′ - GAA ATT AAT ACG ACT CAC
TAT AAA TTT CTA CTA AGT GTA GAA T -3′.
2. crRNA_R (Macrogen): 5′ - NNN NNN NNN NNN NNN
NNN NNA TTC TAC ACT TAG TAG AAA TTT ATA G -3′
(see Note 1).
3. Phusion High-Fidelity DNA Polymerase (Thermo Scientific).
4. 5× Phusion HF Buffer (Thermo Scientific).
5. dNTPs.
6. TOPcloner™ TA-Blunt Kit (Enzynomics).
7. Exprep Plasmid SV (GeneAll).
8. EcoRI-HF (New England Biolabs).
9. NTPs (ATP, UTP, CTP, and GTP).
10. 100 mM MgCl2.
11. T7 RNA polymerase (New England Biolabs).
12. 1× RNAPol reaction buffer (New England Biolabs): 400 mM
Tris–HCl, 60 mM MgCl2, 10 mM DTT, 20 mM spermidine.
13. RNase inhibitor.
14. 1 M DTT.
15. DNase I (New England Biolabs).
16. 10× DNase I reaction buffer (New England Biolabs): 100 mM
Tris–HCl, 25 mM MgCl2, and 5 mM CaCl2.
17. PCR Product Purification Kit (MGmed).](https://image.slidesharecdn.com/24404378-250511200110-8b88bb97/75/Base-Editors-Methods-And-Protocols-1st-Edition-Sangsu-Bae-Beomjong-Song-Editors-48-2048.jpg)