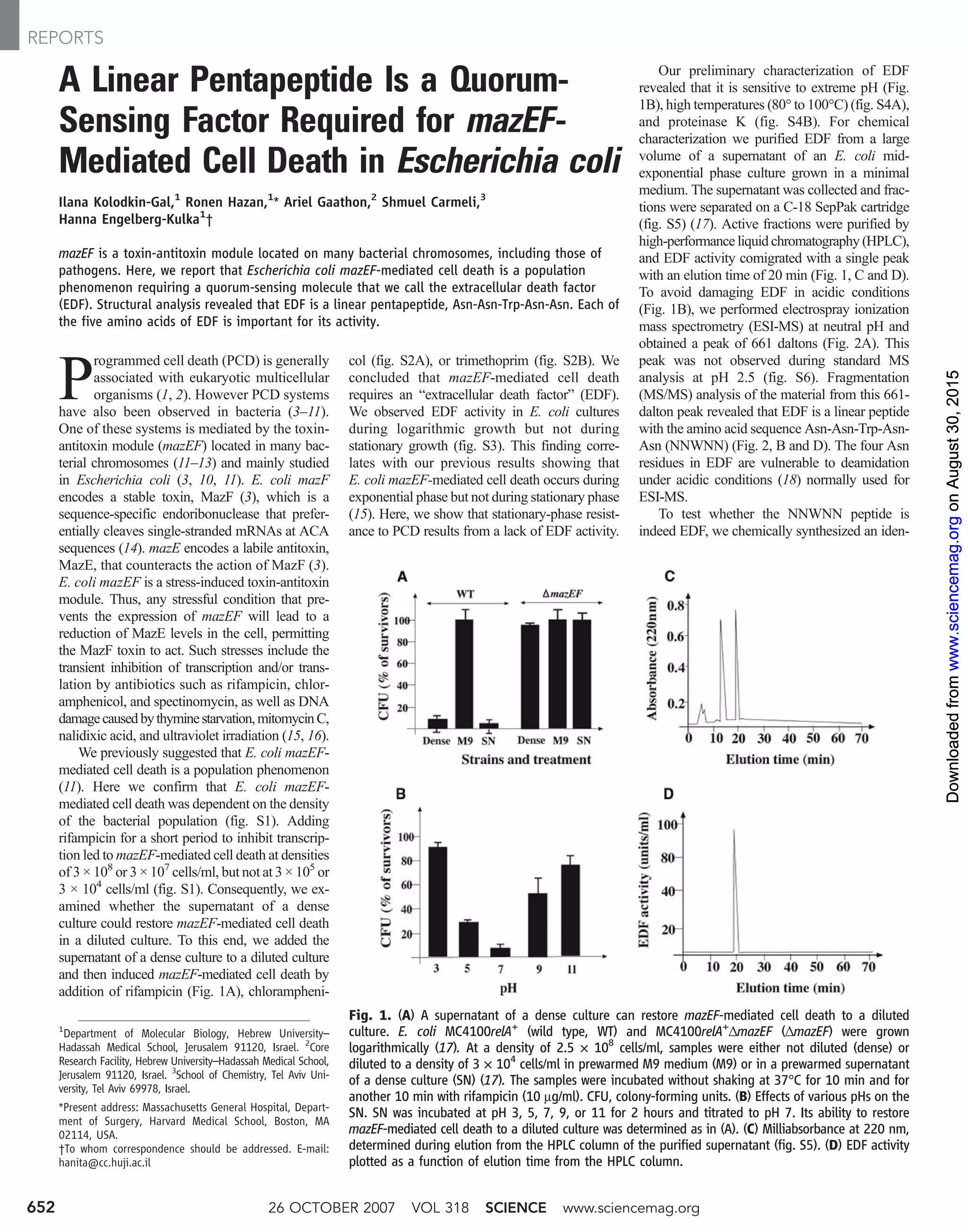
This document summarizes research finding that the mazEF toxin-antitoxin module in Escherichia coli mediates programmed cell death in a density-dependent manner, requiring a quorum-sensing molecule called the extracellular death factor (EDF). Through structural analysis and chemical synthesis, EDF was identified as a linear pentapeptide, Asn-Asn-Trp-Asn-Asn. Mutation analysis showed that each amino acid in the peptide is important for its activity in inducing mazEF-mediated cell death.