1. Hücre Çeperi
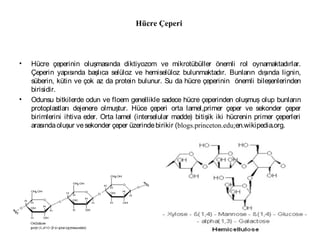
• Hücre çeperinin oluşmasında diktiyozom ve mikrotübüller önemli rol oynamaktadırlar.
Çeperin yapısında başlıca selüloz ve hemiselüloz bulunmaktadır. Bunların dışında lignin,
süberin, kütin ve çok az da protein bulunur. Su da hücre çeperinin önemli bileşenlerinden
birisidir.
• Odunsu bitkilerde odun ve floem genellikle sadece hücre çeperinden oluşmuş olup bunların
protoplastları dejenere olmuştur. Hüce çeperi orta lamel,primer çeper ve sekonder çeper
birimlerini ihtiva eder. Orta lamel (interselular madde) bitişik iki hücrenin primer çeperleri
arasında oluşur ve sekonder çeper üzerinde birikir (blogs.princeton.edu;en.wikipedia.org.
2. Orta Lamel
• Önce ipliksi ve damlamsı oluşumlar halinde olup fragmoplast olarak isimlendirilir. Poligalaktronik
asitin yapıya girmesiyle çeper gelişmeye başlar. Orta lamel pektinler bakımından zengindir. Pektinaz
enzimi ile parçalanır. Odunsu dokularda ligninleşmiştir. Hücrenin uzayarak büyümesi sırasında orta
lamelin primer çeperden ayırt edilmesi imkansızdır. Sekonder çeperi gelişmiş olan trakeid ve liflerde
orta lamel çok fazla incelmiştir. Bunun sonucu olarak iki komşu hücrenin primer çeperleri
arasındaki orta lamel tek bir tabaka halinde görülür. Her üç tabakanın içine fazla lignin girdiği
zaman bu durum daha da belirginleşir. Bu durumda orta lamele bileşik orta lamel denir
(www.sigmaaldrich.com; www.bmrb.wisc.edu)
5. Primer Çeper
• Gelişmekte olan ve canlı hücrelerde bulunur. Hemiselüloz ve çok az pektin bulundurur. Ligninleşme
özelliği gösterebilir. Serin ve hidroksiprolin aminoasitlerini içerir. Alanin ve treonin de bulunur. Protein
miktarı büyüme sırasında çoğalır. Hem yapısal hem de enzimatiktir. Primer çeper önemli oranda su içerir.
Primer çeper,protoplazma ile ilişki halindedir. Primer çeper esnek bir yapıya sahiptir (ro.wikiedia.org;
biologie.uni-hamburg.de groups.molbiosci.northwestern.edu .
10. Sekonder Çeper
• Genellikle etkin olmayan ölü hücrelerde bulunur. Mannoz, galaktoz,ksiloz,arabinoz, yani selülozik
bileşikler ve büyük oranda selüloz ve hemi selüloz içerir.Yine kalloz denilen ve β (1-3) glikozit bağı
taşıyan bir glukoz polimeri de içerir. Sporopolenin maddesi içerir. Lignin, süberin, vaks, kütin,CaCO 3,
silika ve müsilajın yapısına girmesiyle değişikliğe uğrayabilir. Odunsu dokuların hücrelerinde dıştan içe
doğru S1, S2 ve S3 diye üç tabakadan meydana gelir (uky.edu).
11. biologie.uni-hamburg.de; Analysis of TETRAKETIDE α-PYRONE REDUCTASE Function in Arabidopsis thaliana
Reveals a Previously Unknown, but Conserved, Biochemical Pathway in Sporopollenin Monomer Biosynthesis; The Plant Cell
22 (12): 4067-4083 (2010).
22. Hücre Çeperinin Büyümesi
• Hücre çeperi kalınlığına ve yüzeysel olmak
üzere iki şekilde büyümektedir. Kalınlığına
büyümede,çeper maddesi üst üste tabakalar
halinde birikir. Buna apozisyon denir ve
sekonder çeperde gözlenir. Kalınlığına büyüme
ya dıştan hücre lümenine doğru yani
sentripetal veya lümende dışa doğru yani
sentrifugal şekilde olur. Kalınlığına büyüme
homojen veya heterojen olabilir.
23. • Yüzeysel büyümede,mikrofibriller birbirinden ayrılır ve
araya yeni maddeler girer. Bu olaya intussusepsiyon
denir. Yeni maddelerin mikrofibriller arasına
girmesinde oksin,turgor basıncı,protein sentezi ve
solunum olayı rol oynar. Bu tip büyüme ligninleşmiş
primer çeperli ve selülozlu hücrelerde görülür.
• Çeperlerin bazılarında yerel olarak kalınlaşmış bölgeler
vardır. Buna mozaik büyüme denir ve kalburlu
borularda görülür. Multinet büyümede ise yüzey
gerilimi ile mikrofibrillerin düzenlenmesi değişir ve
bunlar bir ağ görünümünü alırlar.
26. Geçitler
• Sekonder çeperlerde derinlik, genişlik ve yapı
bakımından farklılaşmış bölgelere geçit denir.
Primer çeperdeki çukurlara, primer geçit alanı
denir. Geçitler, komşu hücreler arası ilişkiyi
sağlarlar.
27. a) Basit Geçit
• Geçit odasının iki yanı birbirine parelel olarak
düzenlenmiştir.
Şekil 15
(Akman ve ark., 1982)
28. b) Kenarlı Geçitler
• Özellikle trake ve trakeid gibi iletim
demetlerinin yapısında bulunur. Sekonder
çeperde lümene doğru bir kenar oluşmuştur.
Karşıdan bakıldığında ‘V’ şeklinde zar
kalınlaşmasının ortasında kalan bir havuzcuğa
benzer. Çukurluğun geçit kenarı ile örtülen
bölgesine geçit odası denir. Kenardaki
açıklığa,geçit açıklığı adı verilir. Geçit zarı,orta
kısmında kökeni primer hücre zarına zıt olan
torus adlı bir kalınlaşmaya sahiptir.
31. c)Yarı Kenarlı Geçitler
• Bir kenarlı geçitle bir basit geçitin birlikte
oluşturduğu geçit çiftleridir. Bunlarda kenarlı
geçitlerden farklı olarak torus bulunmaz.
Torus, basınç farklılıklarından dolayı geçit
zarının parçalanmasını önlemektedir. Yarı
kenarlı geçitlerde geçitin bir kısmı canlı olduğu
için suyun geçiti parçalaması olanaksızdır.
32. PLAZMODESMATA
• Geçitlerin dışında hücreler arası ilişkiyi sağlayan ince
yapılardır. Plazmodesmata, komşu hücrelerin
protoplastları arasında uzanan çok ince protoplazmik
köprüler olarak tanımlanabilir. Bunlar, canlı hücrelerde
bulunurlar ve hücre öldükten sonra protoplastla
birlikte kaybolurlar. Tohumların çeperleri kalınlaşmış
endosperm hücrelerinden rahatlıkla görülebilir.
Plazmodesmatalar, çoğunlukla geçitlerin bulunduğu
yerlerde toplanmıştır. Geçit bulunmayan hücrelerde
çeper yüzeyine dağılmış gibi görünürler. Bunlar aracılığı
ile besin maddelerinin iletimi sağlanır.
33. Şekil 18. Kalın çeperli endosperma hücrelerinde plasmodesmalar. A. Diospyros (Trabzon
hurması); B, Phoenix (Hurma); C, Phytelephas, ol. orta lamel; pç, primer çeper; ps,
plasmodesmalar (Açıklama metinde), A ve B orijinal olarak; C. HALBSGUTH’a göre
STRASBURGER’den değiştirilerek çizilmiştir.
(Yakar, 1983’ den)
34. DOKU BİLİMİ
• Geniş anlamda hücre topluluklarına doku sistemi denir.
Aynı kökenden gelen,aynı görevi yapan, aynı şekilde
farklılaşmış, birbiri ile sıkı ilişkide olan hücre topluluğuna
doku denir. Hücrelerin bir arada bulunması her zaman doku
oluşumuna neden olmaz. Örneğin: bazı ipliksi alglerde
hücreler yanyana dizilmesine rağmen protoplastlar
arasında metabolik bağ olmadığından doku oluşumu
yoktur. Ascomycetes sınıfından Claviceps purpurea Secale
cereale üzerinde sıkı bir misel kitlesi meydana getirir.
Eczacılıkta drog olarak kullanılan bu kitle gerçek bir doku
olmayıp pseudoparenkima adını alır.
• Gelişmiş bitkilerde dokular bölünür dokular (meristem),
bölünmez dokular (sürekli dokular) diye ikiye ayrılır.
35. Meristemler
• Embriyo gelişiminin ilk safhalarında tüm hücreler
bölünme yeteneğine sahiptirler. Fakat daha sonraki
safhalarda hücre bölünmesi sınırlıdır. Bölünme ileri
safhalarda yalnızca bitkinin özel kısımlarında ve
embriyonik özellikteki dokularda gerçekleşir.
Gelişmekte olan bitkilerdeki bu embriyonik dokulara
meristem denir. Gövde korteksi ve gelişen vaskuler
dokular gibi yapılarda da hücre bölünmesi olabilir.
Fakat bu dokularda bölünme sayısı sınırlıdır.
Meristemler,geçici olarak dinlenme safhasında
bulunabilirler. Bitkinin aktif olduğu zamanlarda bile
bazı koltuk altı tomurcukları uyku halinde olabilirler.
37. MERİSTEMATİK SÜREKLİ
Hücreler ince çeperli Hücreler kalın çeperli
Protoplazma yönünden zengin Protoplazma yönünden fakir
Hücre arası boşluklar yok Hücre arası boşluklar var
Çekirdek hücre hacmine göre Çekirdek hücre hacmine
büyük göre küçük
Diktiyozom ve ER nadir Diktiyozom ve ER çok sayıda
Vakuoller az sayıda Vakuoller çok sayıda
Plastidler nadir Plastidler çok sayıda
Mitokondri sitoplazmanın % 12 Mitokondri sitoplazmanın
sini kaplar. daha az bir kısmını kaplar.
39. • Yüksek bitkilerde döllenme sonucu oluşan zigot bölünerek
embriyoyu verir. Embriyo esas bitkiyi meydana getirmek
için devamlı olarak bölünme yeteneğindedir. Bir bitkinin
hayatında ilk bölünür doku embriyo safhasında
toplanmıştır. Meristematik dokular,ince çeperli,plazması
bol,çekirdeği hücre hacmine göre büyük ve eni boyuna eşit
hücrelerden meydana gelmiştir. Bu dokunun hücreleri
arasında H.A.B. yoktur. Meristematik dokulara ait hücreler
daha sonra sürekli dokulara dönüşürler. Meristemler,
bulundukları yere ve kökenlerine göre sınıflandırılabilrler.
Bulundukları yerlere göre apikal, lateral ve interkalar
meristemler üzere 3 gruba ayrılırlar.
40. • a)Apikal Meristemler: Kök, gövde ve yaprak
uçlarýnda bulunan meristemlerdir
• b)İnterkalar Meristemler: İnternodyumlarda
bulunan meristemlerdir.
• c)Lateral Meristem: Çevreye paralel
bölünmeler ile organların enine büyümesini
sağlayan meristemlerdir. Örneğin:vaskuler
kambiyum ve fellogen
43. • Meristemler kökenlerine göre primer ve
sekonder meristem olmak üzere ikiye ayrılırlar.
Primer meristemler embriyonik hücrelerden
gelişen meristemlerdir. Sekonder meristemler ise
sonradan bölünme yeteneği kazanan
meristemlerdir. Palmiye, muz gibi bazı
monokotiledonlarda gövde kalınlaşması uç
kısımlarda meydana geldiği için kalınlaşmanın
primer yapıda olduğu düşünülür ve bu şekilde
kalınlaşma gösteren meristemlere primer
kalınlaşmış meristem adı verilir.
44. APİKAL MERİSTEMLER
• Apikal meristemlerle ilgili olarak histogen teori ve tunika-korpus teorisi olmak üzere iki
teori vardır. Histogen teoriye göre angiospermlerde büyüme noktasında 3 tabaka vardır.
Bunlar,dıştan içe doğru dermatogen,periblem ve plerom adını alırlar. Dermotogen
epidermisi,periblem korteks ve mezofili,plerom merkezi silindiri meydana getirir. Fakat
Spermatophyta ‘ların çoğunda periblem ve pleromu açıkca ayırmak mümkün olmadığı
için tunika-korpus teorisi geliştirilmiştir. Buna göre büyüme noktasında tunika ve korpus
olmak üzere iki tabaka vardır. Tunika en dış tabaka olup bir kaç sıralıdır ve bölünme
antiklinal yöndedir. Korpus ise merkezi kısmı oluşturmakta olup düzensiz ve büyük
hücrelerden meydana gelmiştir ve bu tabakada bölünme hem periklinal, hem de
antiklinal yöndedir (click4biology.info;content.answcdn.com)
46. • Kökte yaprak bulunmadığı için koruma görevini kök
ucunu yüksük gibi saran kaliptra dokusu yapar.
Kaliptrayı oluşturan hücreler kısa ömürlüdür.
Özellikle dış taraftaki hücreler pelteleşerek kökün
meristem bölgesinin toprak içinde kolayca
ilerlemesini sağlarlar.
Şekil 19. Gövde (A) ve kökün (B) uç kýsýmlarýnýn boyuna kesitini gösteren þema pm.
promeristem d. dermatogen, pb. periblem, pl. plerom, t.tunika, k.korpus, kl. kaliptra,
ys.yaprak sürgünü.
(Özyurt, 1986’ dan)
48. Tunica-Corpus model of the apical meristem (growing tip). The epidermal (L1) and
subepidermal (L2) layers form the outer layers called the tunica [disambiguation needed]. The
inner L3 layer is called the corpus. Cells in the L1 and L2 layers divide in a sideways
fashion, which keeps these layers distinct, whereas the L3 layer divides in a more
random fashion; en.wikipedia.org)
50. Vejetatif Sürgün Ucu Meristemi
• Bu meristem,gövdenin yeni yaprak ve dokularının en uç kısımlarında meydana gelir. (biology.kenyon.edu)
51. Yüksek Bitkilerde Üretken Apikal Meristemi
• Bitkilerin yaprak ve tomurcuklarını veren vejetatif apeksin değişmesiyle meydana gelir. Bu apikal meristem, çiçek ve çiçek durumlarını
verir. Bazı araştırıcılara göre, çiçek sürgünleri ile vejetatif sürgünler homologtur. Bazılarına göre çiçek metamorfoza uğramış bir
sürgündür. Çok sayıda araştırıcıya göre, bitkilerdeki vejetatif apikal meristem zamanla üretken apikal meristeme dönüşür Reproduktif
gövde ucunda çiçek gelişimi sırasında apikal dominans kaybolur sonra DNA sentezinde artış,nukleus çapında artma ve yeni
ribozom meydana gelmesi görülür.
• ; a) Sürgün apikal meristemi b)Çiçek meristemi c) Çiçek tomurcuğu
• Expression analysis of ANTHOCYANINLESS2 gene in Arabidopsis; Plant Science 175 (2008) 853–857
• Hiroyoshi Kubo, Mutsumi Kishi , Koji Goto
52. Kök Ucu Apikal Meristemi
Olgun bir tohum embriyosunda sadece kök promeristemi veya embriyonal radikula
görülmektedir. Bazı eğreltilerde ve at kuyruklarında tüm kök bir tek apikal hücreden gelişir.
Bir çok bitkide lateral ve adventif köklerin promeristemler, primer kök meristemine benzer
(www.infovisual.info)
57. Lateral Meristemler
• Bulundukları organların lateral kısımlarında
bulunurlar. Bu tip meristemler,periklinal
olarak bölünürler. Vasküler kambiyum ve
mantar kambiyumları bu tip meristemlerdir.
59. Tomurcuk ve Kaliptra
• Büyüme noktasındaki genç hücreler,ince çeperli olduğundan dış etkenlerden kolayca zarar
görebilirler. Bu nedenle büyüme noktaları koruma altına alınmıştır. Gövde uçlarında koruma
görevini tomurcuklar üstlenmiştir. Genç yapraklara sıkıca sarılmış büyüme noktalarına
tomurcuk denir. Tomurcukların içinde korunmuş olan uç kısımdaki hücrelerin sürekli bölünme
ve büyümesiyle uzama meydana geldikçe tomurcuğun dış kısmındaki yapraklar açılır ve uç
kısımlarında yeni yaprak sürgünleri oluşur. Böylece, büyüme noktaları sürekli olarak örtülü
kalır (www.cactus-art.biz)
60. Sekonder Meristem
• Bazı sürekli doku hücreleri sonradan bölünme
yeteneği kazanır. Böylece sekonder meristem
oluşur. Ağaç gövdelerini örten mantar
dokusunu meydan getiren fellogen ve kök ve
gövdenin enine büyümesini sağlayan
kambiyum buna örnek olarak verilebilir.
Bunlar,primer meristem hücrelerinden daha
uzundur.
62. Gelişme evrelerine göre ikiye ayrılırlar:
• a)Promeristem: Sürgün apikal meristeminin
merkezi ana hücreleridir. Bunlar,periferal
meristeme öz veya rib meristeme dönüţür.
• b)Kitle, levha ve şerit şeklinde meristemler: Kitle
meristemi embriyo gelişiminin erken safhalarında
faaliyet gösterir. Levha meristemi, levha şeklinde
büyümeye neden olur. Örneğin; epidermis. Şeritsi
meristem yanlız bir yönde gelişir. Sıralar halinde
öz ve kortekste olduğu gibi hücreler gelişir.
Organların uzamasında rol oynar.
63. • Meristemler çok büyük bir nukleusa sahiptir.
Diktiyozom ve ER oldukça nadirdir. Sitoplazmanın
%12’sini mitokondri işgal eder. Vakuol olmamakla
beraber apikal meristemde vakuol bulunabilir.
Bunların plastidleri, proplastid şeklinde olup
granasızdır ve çok az klorofile sahiptir.
64. Vasküler Kambiyum
• Sekonder ksilem ve sekonder floemi meydana
getiren lateral meristemlerdir. Genellikle odunsu
gövdelerde meristematik bir silindir oluşturur.
Periklinal olarak bölünür. İç tarafından meydana
gelen hücreler sekonder ksilemi, dış taraftan
meydana gelen hücreler ise sekonder floemi
meydana getirir. Gövde ve köklerin lateral
kısımlarında yer alır. Bazen yaprak damarlarında
ve petiollerde şerit halinde bulunur. Fasiküler ve
interfasiküler diye iki ţekilde bulunur.
68. • Sitoplazma, ribozomca zengindir. Uzamış veya
izodiyametrik hücreler büyük nukleuslu, ER,
mitokondri ve diktiyozom taşıyan hücrelerdir.
Fusiform inisiyali ve ışın inisiyali diye iki tip
inisiyale sahiptir.
• Fusiform inisiyali sivri uçlu ve uzamış hücrelerden
meydana gelmiştir. Organın uzun eksenine
paralel olarak düzenlenmiştir.
• Işın inisiyali ise izodiyometrik ve oldukça
küçüktür. Merkezden çevreye doğru uzanır ve
floem ve ksilemin öz ışınlarını meydana getirir.
70. • Trake, trakeid, lifler, kalburlu borular, ksilem ve floem
parenkiması fusiform inisiyalden, buna karşın bir
organda yatay düzenlenen vasküler ışın hücreleri ışın
inisiyalinden gelişir. Fusiform hücrelerinin
düzenlenişine göre iki tip kambiyum yapısı vardır.
• a) Tabakalı kambiyum: Bütün fusiform inisiyaller
yatay sıralar şeklinde düzenlenmiştir. Böylece hücre
uçları aynı hizadadır.
• b) Tabakalı olmayan kambiyum: Fusiform
inisiyaller kısmen bir diğerinin üzerine gelir. Yani hücre
sonları aynı düzeyde değildir.
71. Kambiyumda Mevsimsel Değişmeler
• Ilıman bölgelerde kambiyal aktivite
mevsimle,tropik bölgelerde ise su miktarı ile
kontrol edilir. Vasküler kambiyum
sıcaklık,soğuk,kuraklık gibi durumlarda dormant
halde kalır. Tropikal bölgelerde kambiyal aktivite
yoktur veya zayıftır. Ilıman bölgelerde ilkbaharda
oksinin bazipetal hareketi ile mitotik aktivite
kambiyum boyunca yayılır. Sonbaharda ise
kambiyal bölge daralır. Ilkbaharda ise kambiyum
hücrelerin genişlemesi ile ışınsal çeperler zayıflar
ve gövde ve köklerin kabukları kolayca soyulabilir.
72. • Büyüme mevsiminde fusiform hücreler geniş hacimli ve fazla uzamış şekildedir. Uzamış
nukleus merkezde yer alır ve ince bir sitoplazma tabakası ile çevrilidir. Sonbahar odununda
ise fusiform hücreleri dar ve kısadır.Hücrelerde çok vakuol vardır.
• İlkbahar odununda hücreler su alınca çok sayıda vakuol birleşerek büyük bir vakuol oluşturur
ve sitoplazmik akım hızlanır. Sonbahar odununda ise büyük vakuoller, küçük vakuollere
dönüşür ve sitoplazmik akım zayıflar.
• İlkbahar odununda mitokondriler küçük ve küreseldir ve granülsüz ER oranında azalma olur.
Sonbahar odununda ise mitokondriler büyük ve küresel şekillerini kaybetmiştir ve hücreler
granülsüz ER bakımından zengindir.
• İlkbahar odununda diktiyozomlar aktif, sonbahar odununda inaktif durumdadır.
• İlkbahar odununda vakuolün osmotik potansiyeli düşük, sonbahar odunuda ise yüksektir.
• İlkbahar odununda nukleus uzar, sonbahar odunuda ise nukleus kısalır.
• Bazı bitki türlerinde ilkbahar odununda (Acer sp.gibi) kışın nişasta birikimi görülebilir. Ancak
bazı türlerde bunun tersi olur.
73. • İlkbahar ve sonbahar odununda gözlenen bu tip
farklılıklar ya da kambiyumdaki mevsimsel
değişiklikler:
– a) Bitkinin çok yıllık olmasına
– b) oksin hormonunun optimal düzeyde olmasına
– c) Genlerin gerekli enzimleri sentezine
– d) ışık, sıcaklık ve O2 miktarı gibi dış etkilere bağlıdır.
• Genellikle vasküler kambiyum ksilem tarafına
doğru floeme göre daha fazla sayıda hücre verir.
74. Fig. 2. Cambium activity during 2005 growth season. (a) Increased number of cambial (CC) and expanding cells (PC)
give evidence that cell divisions on 25 February already
started. (b) Period of maximal cell production with xylem cells in different phases of cell development; PC, secondary
wall formation (SW) and mature cells (MT) (2 June). (c)
Dormant CC on and mature xylem below it on 26 December. Scale bar 100 lm. Cambial activity, wood formation and
sapling survival of Pinus halepensis exposed to different irrigation regimes
Martin de Luis , Klemen Novak, Jose Raventos , Jozica Gricard, PeterPrislan , Katarina CufarForest Ecology and
Management Volume 262, Issue 8, 15 October 2011, Pages 1630–1638
75. Fig. 3. Structure of different types of tree-rings formed in 2005 in Pinus halepensis. (a) Normal growth ring. (b) E-ring with
latewood like cells in earlywood (arrowhead).
(c) L-ring with earlywood like cells in latewood (arrowhead). Dashed lines represent the boundary between 2004 and 2005 tree
78. Novel aspects of transcriptional regulation in the winter survival and maintenance mechanism of poplar; Tree
Physiology 31:208-225,2011)
79. • Parazitik bitkilerin çoğunda parazit hücreleri
konak vasküler kambiyumunu enfekte ederler.
Bu kambiyuma zarar vermese de sekonder
ksilem ve floem otomatik olarak enfekte olur.
80. Sürekli Dokular
• Büyük vakuollü, az protoplazmalı veya protoplazmasını
tamamen yitirmiş ölü hücrelerden oluşan dokulardır.
Hücre çeperleri meristematik hücrelere göre daha
kalındır. Hücre arası boşluklar yönünden genellikle
zengin dokulardır. Hücre arası boşluklar üç şekilde
oluşmaktadır. Orta lamelin hücre köşelerinden başlayıp
çeperlerin yüzey gerilimleri nedeniyle ayrılması sonucu
şizogen, bazı hücrelerin eriyerek yok olması sonucu
lizigen, komşu hücreler arasındaki büyüme farkı
sonucu az büyüyen hücrelerin çeperlerinin gerilim
sonucu kopmasıyla reksigen hücre arası boşluklar
oluşur. Sürekli dokular şu şekilde sınıflandırılabilir.
81. Primer sürekli dokular
Meritematik orjinlerine göre
Sekonder sürekli dokular
Bunlardan primer sürekli dokular, primer meristemlerin, sekonder sürekli dokular ise
sekonder meristemlerin oluşturduğu dokulardır.
Koruyucu Doku
Parenkima Dokusu
Morfoloji ve fizyolojilerine Destek Doku
göre
Salgı Dokusu
İletim Doku
82. Koruyucu Doku (Örtü Dokusu)
• Organların dışında bulunan ve iç kısmındaki
dokuları kuraklığa,mekanik etkilere karşı
koruyan dokudur. Koruyucu doku,epidermal
sistem ve mantarlaşmış koruyucu doku olmak
üzere iki gruba ayrılır.
83. a) Epidermal Sistem
• Protodermden farklılaşan epidermis ve temel
meristemden farklılaşan bazı epidermis altı
dokulardan oluşur. Stoma,tüy,emergensler de
epidermisin yapısına girerler.
84. • Epiderma: Tüm bitki yüzeyini tek hücre sırasından yapılmış
bir tabaka halinde kaplar. Bazı bitkilerde iki veya daha fazla
tabakadan oluşmuş olabilirler. Bu durumda üst tabakaya
epidermis, alt tabakaya hipodermis denir. Epidermis
hücreleri arasında hücre arası boşluklar genellikle yoktur.
Hücre çeperinde genellikle basit geçitler mevcuttur.
Çoğunlukla küçük lökoplastlar taşıyan epidermis
hücrelerinde kloroplast genelde yoktur. Ancak gölge
bitkileri ışığa çıkarılınca lökoplastlar,kloroplastlara
dönüşebilir. Epidermis hücrelerinin üzeri kutikula ile
örtülüdür. Bu tabaka epidermisin direncini artırdığı gibi su
kaybını önler. Epidermisin başlıca görevi korumak olmakla
beraber,bazı bitkilerde su biriktirmeyi de üzerine alabilir.
87. • Stoma: Epidermis hücreleri arasında hücre arası
boşluklar,genellikle bulunmadığından dış ortam
ile epidermis altındaki dokular arasnıda ki gaz
alış-verişi, stoma adı verilen açıklıklarla sağlanır.
Stoma por denilen bir açıklığa sahip iki stoma
hücresinden yapılmıştır. Porun çevresindeki
hücrelere ‘bekçi hücreleri’ denir. Bunlara bitişik
iki veya daha fazla hücreye ise komşu hücreler
yardımcı hücreler denir. Stoma açıklığındaki
değişmeler,bekçi hücreleriyle komşu hücreler
arasındaki turgor farkına bağlıdır.
88. Epidermisin görevleri:
• a)Su regülasyonu: Carpobrotus edulis yapraklarında ki
gibi balon hücreleri su depo etmektedir. Bitkiye su
alımı ve dışarı su verimesi epidermis hücrelerinin
kontrolündedir.
• b) Güneş ışığına karşı koruma: UV ve kırmızı ötesi
ışınlar bitkilerin çoğu için zararlı olduğundan epidermis
bu tip ışınların bitkiye verebileceği zararları önler.
• c) Diğer organizmalara karşı koruma: Böcek, mantar
gibi organizmaların enfeksiyonları epidermis tarafından
önlenebilir.
• d) Biyolojik olmayan ajanlara karşı koruma: Asit
yağmurları, rüzgar gibi etkilerden bitki epidermis
sayesinde korunur.
89. …
• e) Üreme evresi: Epidermis ve subepidermis
polen tanesinin stigmaya ulaşmasında görev
alır. Yine floral ve ekstra floral nektaryumlar
epidermis tarafından salınır.
• f) Salgı görevi:
90. Stoma komşu hücre ilişkisine göre şu gruplara ayrılır.
• a) Mezogen tip: Bekçi hücreleri ve yardımcı hücreler
aynı ana hücreden meydana gelmiştir. Bu hücrenin
bölünmesiyle önce yardımcı hücreler sonra stoma
hücreleri oluţur.
• b) Perigen tip: Komşu ya da yardımcı hücreler stoma
hücreleri ile aynı ana hücreden meydana gelmemiştir.
Bu şekilde Gramineae tipi stoma meydana gelir.
• c) Mezoperigen tip: Yardımcı hücrelerden bazıları
stoma hücreleri ile ilgilidir. Diğerleri değildir.
91. Factors which cause stomata to open or close
1. Light generally causes stomata to open and darkness to close. In some succulent plants
however, this trend is reversed and the plants open their stomata at night when they can take
advantage of the cooler evening temperatures for gaseous exchange, thus reducing water loss.
2. Wilting or water deficit causes the stomata of a plant to close thus preventing further water
loss from the leaves.
3. An increase in CO2 causes stomatal closure.
4. Increase in temperature causes stomata to open.
(www.botany.uwc.ac.za; biologie.uni-hamburg.de;
www.biologycorner.com; images.tutorvista.com)
93. • Stoma açıklığını etkileyen faktörler, sıcaklık ve nem, ışık,
havanın bulutlu veya açık olması, CO2 yoğunluğu, pH ve
rüzgardır. Sıcaklık transpirasyonu artırdığı için aşırı sıcaklarda
somalar kapanır. Bağıl nem düşük ise yine aynı olay olur.
Düşük CO2 yoğunluğu stoma açılmasına,yüksek CO2 yoğunluğu
kapanmasına neden olur. pH bunun tersi etki gösterir. Açık
havada fotosentez olduğu için stomalar açılır. Stoma sayısı
fotosentez ile ilgili dokularda fazladır. Stomalar en çok
yaprakta bulunur. Gövdede stoma sayısı azdır. Çiçek ve su
bitkilerinde stoma azalmış veya kaybolmuştur. Köklerde ise
stomaya rastlanmaz. Stoma hücrelerinin diğer epidermis
hücrelerinden farkları, sitoplazmalarının bol olması kloroplast
ve nişasta taşımaları ve hücre çeperlerinin bazı bölgelerinde
kalınlaşma göstermeleridir. Amaryllis,Gramine ve Mnium tipi
olmak üzere üç tip stoma vardır.
94. • Amaryllis tip stoma:Epidermis hücrelerine komţu olan
çeperleri ince,pora bakan çeperleri özellikle stoma
hücrelerinin iç ve dış kısmında iki band teşkil etmek üzere
kalındır. Bu kalınlıklar, pora doğru çıkıntılar meydana
getirebilir.
Şekil 20. Amaryllis tip stoma. L. Levkoplast, vç. ventral çeper, dç.dorsal çeper, bh.bekçi hücresi,
a.stoma açýklýðý, n.nukleus, yh. yardýmcý hücre, kl.kloroplast tb. teneffüs boþluðu
(Özyurt, 1986’ dan)
95. • Gramine tip stoma: Gramineae ve Cyperaceae
familyalarında yaygındır. Stoma hücrelerinin geniş olan uç
kısımları ince,aradaki dar kısmı çok kalın çeperlidir.
Şekil 21. Gramine tip stoma A. Yüzeysel B.a-a1 yönünde C. b-b1 yöynünde alýnan kesit
(Özyurt, 1986’ dan)
96. • Mnium tip stoma: Bazı karayosunları ve
eğrelti otlarında rastlanır. Amaryllis tipi
stomaya benzemekle beraber stoma açıklığına
bakan çeperler ince,epiderma hücreleri ile
komşu olan çeperleri kalındır.
98. • Tüyler: Epidermis hücrelerinin dışa doğru meydana
getirdiği uzantılardır. Tüyler, korunma ,
savunma,tırmanma ve salgı gibi görevler yaparlar.
Şekil 22. Çeþitli tipte tüyler A. Kalkan tüyü B. Çok hücreli dallanmýþ bir tüy. C. Kalkan
tüyünün üstten görünüþü. D. Yakýcý tüy. a.Yakýcý b. Basit tüy c.ucu kapak baþ kýsmý E.
Peltat guddeli bir tüy
(Özyurt, 1986’ dan)
99. • Emergensler:Epidermis altındaki dokulardan
köken alan çıkıntılardır. Salgı ve koruma görevi
yaparlar.Örneğin:Rosa ve Rubus sp. ‘deki
dikenler
100. b) Mantarlaşmamış koruyucu doku
• Bu dokuyu meydana getiren hücrelerin
çeperlerine süberin maddesi girmiţtir. Primer
veya sekonder kökenli olabilir.
101. • Primer mantarlaşmış koruyucu doku:
Epidermis veya altındaki dokunun hücre
çeperlerinin mantarlaşmasıyla meydana gelir.
Çoğunlukla hücre arası boşluklar
bulundurmaz. Kök emici tüylerinin
dökülmesiyle görevini yapamayan epidermis
altındaki dokulardan meydana gelen
eksodermis,örnek olarak gösterilebilir.
102. • Sekonder mantarlaşmış koruyucu doku: Dış
etkilerden ötürü veya yaşlanan ve
kalınlaşmaya devam eden odunsu bitkilerin
eksenlerinin kalınlaşmasıyla sürekli bir doku
olan epidermisin gerilmeye dayanamayıp
parçalanması sonucu epidermisin yerini alır.
Bu dokuda epidermisin yerini
periderm,stomaların yerini lentiseller alır.
103. • Periderm: Fellogen, fellem ve felloderma olmak üzere
üç tabakadan oluşur. Fellogen veya mantar kambiyumu
büyük nukleuslu,küçük vakuollü ve fazla sitoplazmalı
hücrelerden oluşmuştur. Fellogen dışa doğru mantar
doku yani fellemi,içe doğru ise fellodermi verir.
Felloderm hücreleri oldukça düzenlidir. Bu hücreler
canlı selüloz çeperlidir. Mantar hücrelerinin çeperleri
ince olmakla beraber çok fazla kalınlaşabilir. Fazla
kalınlaşmış çepere sahip hücrelerden oluşan mantar
dokuya taş mantarı denir. Süberin suya karşı geçirgen
olmadığından mantar doku bitkinin su kaybına engel
olur.
104. • Mantar hücreleri, genellikle hava ile dolu
olduğundan güneş ışınlarını doğrudan
geçiremeyeceğinden bitkiyi fazla ısınmaya karşı
da korur. Mantar oluşumu bazen daha derinlerde
de meydana gelebilir. Örneğin: yaprak dökülmesi
yaprak sapı ile gövde arasındaki mantar dokunun
oluşumu ile olur. Ağaç gövdelerinde kabuk
parçalarının pullar halinde kopup dökülmesi de
çapın artmasıyla mantar dokunun buna ayak
uyduramayıp çatlaması sonucu oluşur.
105. • Yaralanıp ölen hücreler, yüzeyde bir yara zarı
oluştururlar. Bu zarın altındaki hücreler de
süberinleşerek ve ligninleşerek kapama tabakasını
oluştururlar. Bunun altında fellogen meydana gelir. Ölü
yara örtüsü dışarı doğru taşınır. En fazla
Dicotyledones’lerde rastlanır.
Şekil 23. A, B, Pyrus’ta kýsmen epidermis, kýsmen altýnda periderma kökeni, C, D, Prunus’ta ayný durum (Esau’dan)
(Akman ve ark., 1982’ den)
106. Lentisel
• Çoğunlukla ağaçların gövde ve köklerinde kolayca görülebilen
kabarcıklardır. Malus sylvestris’de olduğu gibi meyva
yüzeylerinde de bulunabilirler. Vitis vinifera gibi tırmanıcı
bitkilerde genellikle bulunmamaktadır. Özellikle çok yıllık
bitkilerde stomaların yerini alırlar. Gevşek sıralanmış ve
süberinsiz haldedirler. Hücre arası boşluklar yönünden
zengindirler. Lentiseller stomanın alt kısmında oluşmaya
başlarlar. Gövdenin primer büyümesi sona ermeden ve
periderm meydana gelmeden gelişirler. Bazen de primer
büyümenin sonunda peridermle birlikte meydana gelirler.
Esas oluşum zamanı organın ilk büyüme mevsimidir. Organ
uzaması tamamlanmadan önce de oluşabilir. Bitki türlerinde
her stoma altında veya her stoma grubunun altında bir
lentisel bulunur. Genelde düzensiz bir dağılım gösterirler.
Sonbaharda mantar dokusu lentisellerin kapanmasını
sağlamaktadır.
107. Şekil 24. Sambucus dalýndan alýnan enine kesitlerde lentiseller A. Genç B. Yaþlý faz e.
epidermis, p.mantar, pg. fellogen, pd.felloderm, k.komplementer hücreler
(Özyurt, 1986’ dan)
108. Parenkima
• İnce çeperli,çokgen şeklinde canlı ve fizyolojik yönden
değişiklik gösteren hücrelerden oluşmuş bir dokudur.
Bitkinin her tarafında bulunduğu için temel doku adını
alır. Özellikle vasküler dokular bu yapı içine gömülüdür.
Tüm apikal meristemler ve üreme hücreleri
parenkimatik yapıdadır. Yaralanan bölgelerde bu doku
hemen meristematik özellik kazanır.
• Gövde ve kökün öz ve korteksinde, yaprak mezofilinde,
sukkulent meyvelerin etli kısımlarında ve tohum
endospermlerinde bulunurlar. Ayrıca özel olarak ksilem
ve floem parenkimaları da vardır.
109. • Parenkima hücreleri genellikle polihedral ve
izodiyametrik olmalarına rağmen bazen uzamış ve sivri
uçlu olabilirler. Bu durumda bunlara prosenkima denir.
Yaprak mezofilinde ve diğer bazı bitki kısımlarında
loblu ve katlanmış ve kollu palizat hücreleride görülür.
Örneğin; Lilium sp.’de loblu, Pinus sp’de kollu ve
Juncus sp’de yıldız şeklinde parenkima vardır.
• Parenkima hücreleri genelde selülozik ve primer
çeperlidir. Ancak bunlar kalınlaşıp depo görevi de
yapabilirler. Genelde depo edilen madde
hemiselülozdur. Bu ţekilde hücrelere genellikle Coffea
arabica, Phoenix dactylifera, Asparagus sp’in
endospermlerinde rastlanır.
111. • Parenkima hücreleri vakuol öz suyunda serbest olarak
veya sitoplazmada katı halde birçok madde depo
ederler. Örneğin Solanum tuberosum yumrularında ve
rizom tipi gövdelerde vakuol öz suyunda proteinler;
sitoplazmada da nişasta taneleri bulunur.
• Parenkima hücreleri genellikle tanen de bulundurur.
Bunlar,ya yapraklarda olduğu gibi devamlı bir tabaka
oluştururlar veya gövdedeki gibi ayrı ayrı bulunur.
Tanenli hücreler genellikle özün en dış kısmında
bulunur. Parenkima hücrelerinde mineral maddeler de
depo edilerek kristaller oluşturulur.
112. • Olgun parenkima hücreleri ya Poaceae
endospermlerindeki gibi sıralar halindedir
veya meyve ve sukkulent organlardaki gibi
hücre arası boşluklar yönünden zengin
parenkima bulunur. Karanlıkta gelişen kök öz
gibi dokularda hücre arası boşluklar yönünden
zengin parenkima bulunur.
• Parenkima dokusu görevlerine göre çeşitli
tiplere ayrılır.
114. • 1) Asimilasyon parenkiması: Fazla miktarda
kloraplast ve lökoplast bulundurur ve
klorenkima adını alır. En belirgin şekilde
yaprak mezofilinde görülür.
• 2) Depo parenkiması: Organik maddelerin
depo edildiği parenkima tipidir.
116. • 3) Su depo parenkiması: Aloe sp., Agave sp.,
Opuntia ficus indica,Carpobrotus edulis ve
Meserbryantahemum sp. gibi bitkilerin
fotosentez organlarında klorofilsiz su dolu
parenkima görülür. Bu dokuyu oluşturan hücreler
genellikle büyük, ince çeperli hücrelerden
meydana gelmiştir. Bu hücreler ya düzenli sıralar
halindedir veya palizat parenkiması hücreleri gibi
uzamışlardır. Her hücrenin ince bir tabaka halinde
sitoplazması,nukleusu ve musilajlı geniş bir
vakuolu vardır.
117. Su depo parenkiması; Slightly collapsed water-storage parenchyma cells in the inner cortex of
a Brasilicereus plant that had not been watered for several weeks;www.sbs.utexas.edu)
118. • 4) İletim parenkiması: Öncelikle karbonhidratların
taşınmasını sağlayan parenkimalardır.
• 5) Aerenkima: Bataklık ve su bitkilerinde bulunan ve
bol miktarda gaz depo eden parenkimadır.
Havalandırma parenkiması adını alır. Hem gaz alış-
verişi, hem de yüzmeyi sağlar(Carpobrotus sp. gibi).
Bataklık ve su bitkilerinde bulunana parenkima ise
hava depo etmekte olup aerenkima adını alır.
Parenkima hücreleri,bol miktarda tanen ve kristal depo
ederler.Parenkima hücreleri arasında hücre arası
boşluklar bulunabilir veya bulunmayabilir.
121. DESTEK DOKUSU
• Bitki yapısına belirli bir dayanıklılık ve direnç
kazandıran dokulardır. Bu dokular kollenkima
ve sklerenkima olmak üzere iki gruba ayrılırlar.
Bunların ikisine birden sterom adı verilir
122. Kollenkima
• Kollenkima terimi,hücre çeperinin kalın ve parlak oluşunu
ifade eder. Kollenkima,az-çok uzamış,ligninleşmiş,kalın
primer çeperli hücrelerden meydana gelen canlı bir
dokudur. Morfolojik olarak birbirine benzeyen hücrelerden
meydana geldiğinden homojendir. Genellikle büyümekte
olan gövdelerde, çiçek ve yapraklarda yer alır. Hücre çeperi
fazla gerilme ve esneme özelliğine sahiptir. Bu bakımdan
büyümekte olan organlarda meydana gelen değişikliklere
uyum sağlayabilir. Kollenkima hücreleri izodiyametrik
olabildiği gibi lif gibi uzamış da olabilir. Kollenkima dokusu
kök korteksinde bulunur. Özellikle ışık gören kısımlarda
çok daha iyi gelişir. Gövde ve yapraklarda çevresel olarak
meydana geldiğinden eğilmeye karşı bitkiye dayanıklılık
sağlarlar.
123. • Kollenkima dokusu epidermisin altında meydana gelir.
Yapraklarda ise damarların ya bir tarafında ya da her iki
tarafında bulunur. Bitkinin fazla rüzgar gören kısımları
kollenkima yönünden zengindir. Kollenkima hücrelerinin
çeperleri selüloz ve pektinli maddelerden meydana gelmiştir.
Nadir de olsa kollenkima hücreleri meristematik özellik
kazanabilirler. Kollenkima çeperlerinde lignin bulunmaz ve
basit geçitlidir. Kollenkima dokusu köşe,levha ve lakün
kollenkiması olmak üzere üç gruba ayrılır. Köşe
kollenkimasında kalınlaşmalar hücre köşelerinde meydana
gelir. Levha kollenkimasında kalınlaşma, hücrenin teğetsel
çeperlerinde, lakün kollenkimasında ise kalınlaşma hücre arası
boşluklara bakan çeperlerde meydana gelir. Kollenkima
dokusunda H.A.B. oldukça zengindir. Kollenkima çeperleri, K+
ve Cl- yönünden zengindir. Agglutunin olduğundan
antibakteriyel etkisi vardır. Ancak mantar çeperlerinin
pektinlerini hidroliz ederler.
124. Sklerenkima
• Sklerenkima terimi,çeperin sertliğini ifade eder.
Sklerenkima hücreleri büyüme yeteneklerini
kaybetmişlerdir. Gelişmiş organlarda bulunur.
Kollenkimadan çeper yapısının sertliği,ligninleşmesi ve az su
bulundurmasıyla ayrılır. Ayrıca sklerenkima elastiki
sekonder çeperlidir. Halbuki kollerankima plastik primer
çepere sahiptir. Sklerenkimayı meydana getiren hücrelerin
çeperleri olgun halde hem kalın,hem de odunlaşmıştır.
Çeper kalınlaşması her yerde aynıdır. Olgun fazda hücreleri
kollenkimadan farklı olarak protoplazma içermez. Diğer bir
ifadeyle olgun fazda ölüdürler. Sklerenkima dokusunda
bulunan hücreler, sklerenkima hücreleri (sklerid) ve
sklerenkima lifleri (fiberler) olmak üzere 2 gruba ayrılırlar.
Genel olarak sklerenkima lifleri uzun, sklerenkima hücreleri
ise kısa hücreleridir. Fakat bazen bunun tersi de olabilir.
Sklereidler genellikle parenkima hücrelerinden lifler de
meristematik hücrelerden gelişirler.
125. • Şekil 25. Kollenkima tipleri. A. Sambucus gövdesinde plak kollenkiması. B.
Begonia gövdesinde köþe kollenkiması. C. Lactuca gövdesinde lakün
kollenkiması in.interselüler alan. (Özyurt, 1986’ dan)
126. Sklerankima Hücreleri
• Bunların,genellikle eni boyuna eşittir. Tek veya
gruplar halinde bulunurlar. Camellia ,
Thea,Nymphaea gibi bitkilerin yapraklarındaki
parenkimatik hücreler arasında bulunan kalın
çeperli sklereidlerle idioplast denir. Armut veya
ayvada olduğu gibi meyvelerin etli kısımlarında da
bulunurlar. Fındık ve cevizde olduğu gibi
perikarpta da bulunabilirler. Bunlar şekillerine
göre brakisklereidler (taş hücreleri),
makrosklereidler, osteoskleridler ve
astrosklereidler olmak üzere 4 gruba ayrılırlar.
128. • Şekil 26. Sklereid tipleri a.Astrosklereid b. Brakisklereid c. Makrosklereid d.
Osteosklereid. (Özyurt, 1986’ dan)
129. Sklerenkimatik Lifler (Fiberler)
• Boyu eninden oldukça uzun ve sivri uçlu
hücrelerdir. Çeperleri,genellikle ligninleşmiş
olmakla beraber selüloz çeperli olanlar da vardır.
Genellikle ölü hücrelerdir. Morfolojik olarak
ksilem lifleri ve ksilem dışı lifler olmak üzere iki
gruba ayrılırlar. Ksilem lifleri,indirgenmiş
trakeidlerdir. Libiform lifleri, lif trakeidleri ve
jelatinli lifler (musilajlı) diye üç gruba ayrılırlar. Lif
trakeidleri, libriform liflerden biraz daha uzundur.
Bazen libriform lifleri ve lif trakeidleri bölmelidir
(septalı). Böyle liflere septat lifler denilir. Septat
liflerde fazla miktarda ergastik madde depo edilir.
130. • Ksilem dışı lifler, floemde, iletim demetlerinin etrafında ve korteksde bulunurlar. Bunların
çeperleri çok fazla kalınlaşmışlardır. Şekilleri uzun ve uçları küttür. Basit geçitlere sahiptirler.
Cannabis, Agave, Yucca, Crochorus ve Linum sp. ’de olduğu gibi çok uzun olabilirler.
Bunların ekonomik önemi büyüktür. Jelatinli (musilajlı)liflerde sekonder çeperin en iç
tabakası çol miktarda lignin hemiselüloz bulundurur. Çok su emen bu tabakaya G tabakası
denir ve bu tabaka S3 ‘te bulunup kuruyunca büzülür (www.uriedu; www.aspca.org)
131. Tension wood of sweetgum ( Liquidambar styracifl ua ). (A) Transition region between normal wood (NW, top) and tension wood (TW,
bottom).Note the pink-red color of the gelatinous layer in the G-fi bers in the bottom two-thirds of the image and the marked lack of this
color in the normal wood at the top of the fi gure. Area in box is shown at higher magnifi cation in C. (B) An area of tension wood
showing the large number of G-fi ber cells. The walls of the vessel elements and the ray cells stain a blue-purple, with no trace of pink-red
characteristic of G-layers. Also, vessel elements are much smaller and less frequent than in the normal wood in A. Area in box is shown at
higher magnifi cation in D. (C) Higher magnifi cation of the region in the black box in A showing details of the transition zone (TZ)
between normal wood (left) and tension wood (right). The middle lamella is much thicker in the normal wood than in the TZ cells or the
tension wood. The cells of the transition zone have a thickened, blue-stained secondary wall, but no pink-red stained material is present in
these cells. The blue layer is thinner in the fi bers of the G-layer, and a pink-red layer is internal to the blue layer. (D) The same magnifi
cation as (C), but showing a region of G-fi bers well away from the normal wood and the transition zone. The pink-red G-layer is very
prominent and thick in these cells. The light blue S1/S2 layer is thin, and the dark-purple middle lamella (ML) is also thin. Scale bars: A
and B, 100 μ m; C and D, 25 μ m. IMMUNOCYTOCHEMICAL CHARACTERIZATION OF TENSION
WOOD: GELATINOUS FIBERS CONTAIN MORE THAN JUST CELLULOSE Andrew J. Bowling and Kevin C. Vaughn American
Journal of Botany 95(6): 655–663. 2008.
132. • Şekil 27. Sklerenkima lifi. A. Cinchona korteksinde sklerenkimatik lif B.
Aristolachia’da sekonder ksilemde bölmeli lif g.geçit.(Özyurt, 1986’ dan)
133. İletim Dokusu
Ksilem
• Su ve suda çözünmüş maddeleri ileten dokulardır. Trake, trakeid,
parenkima ve sklerenkima hücrelerinden oluşur. Ksilem dokusu,
bitkinin embriyonik fazı ve bunu takip eden fazlarda farklılık
gösterir. Yani primer ksilem ve sekonder ksilem oluşur. Primer
ksilem proksilem ve metaksilem diye iki gruba ayrılır.
Protoksilemde bulunan trake ve trakeidler, komşu hücrelerin
uzamasına ayak uyduramadıklarından gerilir ve parçalanırlar. Bunlar
primer vasküler sistemde bulunurlar ve bulundukları organın
uzaması tamamlanınca yerini metaksileme bırakırlar. Metaksilem,
organın büyümesi, sona erdikten sonra oluştuğu için bunun
elementleri fazla değişime uğramaz. Vasküler kambiyumun
meydana getirdiği sekonder dokuya ise sekonder ksilem veya odun
denir. Sekonder ksilem,primer ksilemden farklı olarak vertikal ve
horizontal sistemlerden oluşur. Vertikal sistem trake, trakeid, lif ve
parenkima hücrelerinden, horizontal sistem ise parenkima
hücrelerinden meydana gelmiş olup öz ışınlarını meydana getirir.
138. Ksilem Işınları
• Ksilemlerde ışınsal olarak uzanan şeritsi bir
dokudur. Kambiyumdan kortekse doğru
uzanır. Vasküler ışın adını da alır. Gelişimlerine
kambiyumdan başlar ve kambiyumun
faaliyetiyle uzamalarını sonsuz olarak
arttırırlar.
140. Trakeidler
• Cansız, uzun, dar ve iki ucu kapalı, sivri, olgun
fazda protoplastsız ve ligninleşmş hücrelerdir.
Yalnızca kenarlı geçitleri olan deliksiz
hücrelerdir. Delik olmadığından madde alış-
verişi ince geçit zarlarıyla olmaktadır.
Gymnospermae odununda yalnızca trakeid
bulunur.
143. Trakeler
• Trakeidler gibi ölü, ligninleşmiş, kalın çeperli,
enleri geniş, uçları açık, trakeidlere göre daha
kısa boylu üst üste gelmiş ve aralarındaki
bölme zarları erimiş hücrelerdir. Bunlar delikli
(perforat) tüplerdir. Bunlar sayesinde serbest
akım sağlanır. Delikler genelde uç çeperlerde
meydana gelirler. Çeprin delik ihtva eden
alanına perforasyon tablası denir. Tek delikli
olursa uniperforat, çok delikli olursa
multiperforat adını alır.
145. • Şekil 28. Farklı bitkilerin gövdesinden alınmış boyuna kesitlerde farklı tipte
trakeler. A. Acer (Akçaağaç); B. Malus pumila (Yabanielma); C.
Liriodendron (Lâleağacı); D. Betula (Huş-ağacı). (Yakar, 1983’ den)
151. Ksilem Lifleri
• Libiform lifleri ve lif trakeidleri diye ikiye
ayrılır. Libriform liflerinde basit geçitler varken
lif trakeidlerinde kenarlı geçitler bulunur.
153. Ksilem Parenkiması
• Ksilem parenkiması ve ışın parenkiması diye ikiye ayrılır (sols.unlv.edu;
www.puc.edu).
154. Yumuşak ve Sert Odun
• Trake,trakeid ve lif hücreleri fizyolojik
faaliyelerinden önce proplastlatını
kaybederler. Parenkima ise proplastını korur.
Canlı ve aktif iletme işlemine sahip oduna
yumuşak odun,aktif olmayan ve protoplastını
kaybeden oduna da sert odun adı verilir. Bir
ağacın yumuşak odunu iletim, destek, ve gıda
depo etme görevini yapar. Sert odun ise
destek görevini yapar.
155. Yumuşak odun sert oduna dönüşürken
şu değişmeler olur.
• Bütün canlı hücreler protoplastlarını kaybederler.
• Hücre öz suyu çekilir.
• Hücre çeperi yapısındaki su miktarı azalır.
• Canlı hücrelerdeki besin maddesi azalır.
• Tilosis olur.
• Parenkima hücrelerinin çeperleri daha fazla
ligninleşir.
• Hücrede tanen,reçine,yağ,aromatik ve renkli
maddeler birikir.
• Geçitlerin torusları geçit ağzını kapatır.
156. • Bir ağacın yumuşak ve sert odun miktarı ağacın türüne
ve büyüme şartlarına göre değişir. Abies sp., Picea sp.,
Salix sp. ve Populus sp. gibi bitkilerde bariz farklılaşmış
bir sert odun yoktur. Robina sp. Morus sp.ve Taxus
sp.’de ise yumuşak odun miktarı azdır. Eucalyptus sp.,
Acer sp., Castanea sp., Buxus sp. ve Fraxinus sp.’de
odun çok serttir.
• Sert odun,yumuşak oduna göre bakteri ve mantar
enfeksiyonlarına karşı daha dayanıklıdır. Reçine, tanen,
yağ gibi maddelerin birikimi, tilosis ile su iletiminin
durması, protoplastın ortadan kalkması da dayanıklılığı
arttıran etkenlerdir.
157. • Genç ağaçlarda ve yaşlı ağaçların çok çabuk büyüyen
genç kısımlarında sert odun ya çok azdır veya hiç
yoktur. Yavaş büyüyen zayıf ve yaşlı ağaçların gövde ve
köklerinde sert odun miktarı daha çoktur.
• Quercus sp.,Ulmus sp.,Fraxinus sp. gibi ağaçların sene
halkalarında ilkbahar odunu, sonbahar odunundan
daha geniş ve daha fazla sayıda trake ihtiva eder. Bu tip
ağaçların odunlarına halkalı-delikli odun denir.
• Acer sp., Betula sp., Populus sp. gibi bazı ağaçların
odunlarında ise trakeler bütün sene halkalarında
homojen dağılırlar. Bu tip odunlara da dağınık-delikli
odunlar denir.
161. Odun Özü ve Odun Kalbi
• Odundaki büyüme halkaları,5-10 yıl ya da daha fazla
yaşlandığı zaman iletim yeteneğini kaybeder. İç
kısmında oluşan ve iletim yeteneğini kaybeden
halkalara odun kalbi,dışta iletim yapan tabakalara ise
odun özü denir. Her yıl odun özü odun kalbine
dönüşür. Fakat kambiyum yeniden odun özünü
meydana getirdiğinden azalma telafi edilir. Odun
kalbinde lignin ve fenoller biriktiğinden renk koyudur.
Parenkima hücreleri öldüğünden odun özünden daha
dayanıklıdır. Populus, Salix, Abies’de bu ayrım yoktur.
Morus’ta ince Acer, Fagus ve Fraxinus‘ta kalın odun
özü vardır. Ksilemin vertikal sistemi iki tip parenkima
içerir. Apotrakeal parenkimada parenkimanın
trakelerle ilişkisi yoktur. Paratrakeal parenkima da ise
trakelerle yakından ilişkilidir ve trakelere doğru uzanan
uzantılara sahiptir.
163. Gymnospermae Odunu
• Basit ve homojen bir yapıya sahiptir. Trake ve
libriform lifleri yoktur. Parenkima miktarı da
azdır. En fazla farklılaşmış ksilem ışınlarına
Gymnospermae odununda rastlanır.
Gymnospermae odununda ışınlar parenkima ve
trakeidlerden oluşmuştur. Işınsal parenkima
hücreleri asimilasyon maddelerinin iletilmesinde
rol oynar. Bu tip odunlarda yaralanma şizogen ve
lizigen tipteki reçine kanalları genişler ve
nişastanın parçalanması ile içlerinde sakızımsı bir
madde oluşur. Bu olaya gummosis denir.
Sakızımsı madde bitkiyi enfeksiyonlara karşı
korur.
164. Angiospermae Odunu
• Çok kompleks bir yapıya sahiptir. Ksilem
ışınlarının düzenlenişi de değişiktir. Odun
ışınları yalnızca parenkima hücrelerinden
oluşmuştur.
166. Tilosis
• Çok sayıda bitkide ksilem ve ışın parenkiması
hücreleri trake ve trakeid yaralandığı zaman
iletim borularının içersine doğru girerek torba
şeklinde genişler ve böylece su ve inorganik
madde akımı olur. Bu olaya tilosis adı verilir.
Tilosisin gelişimi parenkima hücreleri ile trake
ve trakeidler arasındaki geçitlerin kapatma
zarlarından meydana gelir. Böylece iletim
sisteminin inaktif hale gelmesini sağlayan bu
olay diğer elemanlara geçmiş olur.
171. Floem
• Kalburlu hücreler Gymnospermae’lerde bulunmaz
olgunlaşmış kalburlu borular ER,diktiyozom ve ribozom
içerir. Ayrıca akıcı nitelikte proteinler taşırlar. Proteini
yaralanma sırasında floemde iletimi durdurur.
• Kalburlu hücreler, ilkel yapılı bitkilerde,
Gymnospermae’lerde,Angiospermae’lerde bulunur.
Bunlar ince selüloz çeperli uzun canlı hücrelerdir.
Olgun fazda fazla nukleus bulundurmazlar.
Meristematik fazda ise bol protoplazma ve nukleus
bulundururlar. Kalburlu alanlarda çeperler çukurlaşmış
ve delikler meydana gelmiştir. Bu deliklerden
plazmodesmataya benzeyen sitoplazmik iplikler uzanır.
Birleştirme kolları denilen bu uzantılar kalloz denilen
bir polisakkaritten oluşmuş kallus örtüsü ile
kaplanmıştır. Bu alanlara kalburlu plak denir. Kallus
miktarı,yaşla birlikte artmaktadır.
174. Arkadaş Hücreleri
• Kalburlu boru elementlerinin yanında bulunana
parenkimatik, küçük, dar ve dikdörtgen şeklindeki
hücrelerdir. Kalburlu boru elementi ile aynı
meristematik hücrelerden gelişirler.
Gymnospermae odununda bulunmazlar. Kalburlu
boru elementleri ve arkadaş hücreleri fizyolojik
olarak birbirleriyle ilişkilidir. Kalburlu borular gibi
P-proteinleri bulundurur. Kloroplast ya da
lökoplast, mitokondri, ER ve diktiyozom
bakımından zengindir.
175. Plant Physiology; 3rd Edition; Lincoln Taiz and Eduardo Zeiger
Sinuaer Associates Inc. , In: Chapter 10 p. 195)
177. Floem Parenkiması
• Bol miktarda ergastik madde depo ederler.
Vertikal sistemin parenkimasına floem
parenkiması,horizontal parenkimaya ise floem
ışınları denir. Bir çok bitkide fellogen,floem
parenkiması ve ışınlardan meydana gelir.
178. Floem Lifleri
• Çeperleri basit geçitli ve ligninleşmiş liflerdir. Bölmeli ve musilajlı da olabilirler
• Primer floem,protofloem ve meta floem olmak üzere ikiye ayrılırlar. Protofloemde arkadaş hücreleri bulunmaz ve bu floem
tipi organ büyümesini tamamlanmadan meydana gelir. Hücrelerin nukleussuzdur. Organ büyümesini tamamladıktan sonra
oluşan floeme ise metafloem denir. Metafloem kalburlu elementlerin protofloemden daha uzun ve daha geniştir. Metafloemde
çoğunlukla lif bulunmaz. (6: Floem lifleri www.cmjournal.org)
179. Sekonder Floem
• Sekonder büyüme gösteren bitkilerde oluşur. Sekonder
floemin primer floemden farkları sekonder floemde
ışınsal sıralardaki hücrelerin düzgün dizilişi, kalburlu
boruların organın fazla ve daha geniş oluşu ve liflerin
boylarının daha kısa oluşudur.
• Sekonder floemin vertikal sistemi
Gymnospermae’lerde kalburlu hücreler,
Angiospermae’lerde ise kalburlu borular ve arkadaş
hücreleri oluşturur horizontal sistem ise ışın parenkima
hücrelerinden meydana gelmiştir. Sıcaklık, ışık, asit ve
hidrojen siyanür gibi ket vurucular ve bitkisel
hormonlar floemde madde taşınmasını engelllerler.
180. Sieve tube elements found in angiosperms
1. Some sieve areas are differentiated into sieve plates;
individual sieve tube elements are joined together into a
sieve tube.
2. Sieve plate pores are open channels.
3. P-protein is present in all dicots and many monocots.
4. Companion cells are sources of ATP and perhaps other
compounds and, in some species, are transfer cells or
intermediary cells.
Sieve cells found in gymnosperms
1. There are no sieve plates; all sieve areas are similar.
2. Pores in sieve areas appear blocked with membranes
3. There is no P-protein.
4. Albuminous cells sometimes function as companion cells.
(Plant Physiology; 3rd Edition; Lincoln Taiz and Eduardo Zeiger
Sinuaer Associates Inc. , In: Chapter 10 p. 196)
181. Plant Physiology; 3rd Edition; Lincoln Taiz and Eduardo Zeiger
Sinuaer Associates Inc. , In: Chapter 10 p. 211)
182. Plant Physiology; 3rd Edition; Lincoln Taiz and Eduardo Zeiger
Sinuaer Associates Inc. , In: Chapter 10 p. 201)
183. ATP-dependent sucrose transport in sieve element loading. In the cotransport model of sucrose loading
into the symplast of the sieve element–companion cell complex, the plasma membrane ATPase pumps protons out
of the cell into the apoplast, establishing a high proton concentration there. The energy in this proton gradient is
then used to drive the transport of sucrose into the symplast of the sieve element–companion cell complex through
a sucrose H+ transporter Plant Physiology; 3rd Edition; Lincoln Taiz and Eduardo Zeiger
Sinuaer Associates Inc. , In: Chapter 10 p. 209)
184. TABLE 10.1
Characteristics of the two types of sieve elements in
seed plants
Sieve tube elements found in angiosperms
1. Some sieve areas are differentiated into sieve plates;
individual sieve tube elements are joined together into a
sieve tube.
2. Sieve plate pores are open channels.
3. P-protein is present in all dicots and many monocots.
4. Companion cells are sources of ATP and perhaps other
compounds and, in some species, are transfer cells or
intermediary cells.
Sieve cells found in gymnosperms
1. There are no sieve plates; all sieve areas are similar.
2. Pores in sieve areas appear blocked with membranes
3. There is no P-protein.
4. Albuminous cells sometimes function as companion cells
Plant Physiology; 3rd Edition; Lincoln Taiz and Eduardo Zeiger
Sinuaer Associates Inc. , In: Chapter 10 p. 196)
185. Gymnospermae Floemi
• Basit yapılıdır. Arkadaş hücreleri yerine
albuminli hücreler vardır. Işınlar genellikle tek
sıralıdır (www.studyblue.com).
186. Dikotiledon Floemi
• Vertikal sistem,arkadaş hücreleri ve kalburlu boru hücrelerinden meydana gelir. Vertikal sistem lifleri
birçok bitkide teğetsel kollar halindedir (www.studyblue.com).
189. Aktif Olmayan Floem
• Yeni büyüme mevsimi başlamadan önce bütün
kalburlu borular aktif değildir. Bu tip
floem,genellikle canlı parenkima dokusunu
korumaktadır. Aktif olmayan floem miktarı
fellogen faaaliyetine bağlıdır. Fellogen
derindeki dokulara gitmemişse bu tip
bitkilerde geniş bir aktif olmayan floem
bölgesi vardır.
190. Sekonder Floemin Ekonomik Önemi
• Malvaceae, Moraceae, Tiliaceae sekonder
floeminden elde edilen lifler ekonomik önem
taşır. Floemden baharat, kinin gibi maddeler
elde edilir. Ayrıca reçine gibi maddelerin de
kaynağıdır.
191. Vasküler demetler
• Bitkilerin vasküler sisteminin serbest olan
kollarına denir. Prokambiyumdan oluşur.
Vasküler demetler, kolletral, bikollateral,
konsantrik ve radyal olmak üzere 4 tiptir.
192. • I) Kollateral Demetler: Bu tip demetlerde kambiyum varsa
açık kollateral demet,yoksa kapalı kollateral demet adını
alır. Kapalı kollateral demet monokotillerde,açık kollateral
demet Gymnospermae’lerde görülür.
• II) Bikollateral Demetler: Bu tip demetlerde ksilem iki
floem tabakası arasında yer alır. Cucurbitaceae’ de görülür.
• III) Konsantrik Demetler: Hadrosentrik demet eğreltilerde
ve çiçek organlarında görülür. Floem dışta,ksilem içtedir.
Leptosentrik demette floem içte,ksilem dışdadır.
• IV) Radyal Demetler: Kibrit otlarında görülür. Kökün primer
yapısında da rastlanır. Yıldız şeklindedir.
193. • Şekil 30. Çeşitli iletim demetleri, a. Kollateral kapalı demet; b. Kollateral
açık demet; c. bikollateral açık demet; d. leptosentrik demet; e.
hadrosantrik demet; f. radyal demet, k. kambiyum, p. parankima, Ksilem
siyah, floem noktalı gösterilmiştir. (Özyurt, 1986’ dan)
194. Salgı Dokusu
• Zamk, reçine, eterik yağlar, su, nektar, musilaj,
kauçuk, alkoloid ve benzer maddelerin
salınmasından sorumlu dokudur. İki grupta
incelemek mümkündür.
– 1) Hücre içi salgılar
– 2) Hücre dışı salgılar
195. • Salgıların meydana geldiği yer daima hücre
plazmasıdır. Bunlar, hücre öz suyunda veya
vakuollerde de toplanabilir. Bu iki durumda da
salgılanan madde hücre içinde kalır. Bu
şekildeki salgılara intraselüler, hücre dışına
verilen salgılara da ekstraselüler salgılar denir.
196. Hücre İçi Salgılar
• Epidermisin altında veya daha iç dokularda
meydana gelirler.
– 1. Salgı hücreleri
– Salgı maddesini kendi içlerinde saklarlar. Salgı
maddesi Ca-oksalat kristallerinde olduğu gibi
hücre içinde katı halde bulunurlar. Bazen salgı
maddeleri eterik yağlar gibi sıvı halde de olabilir.
Buna en tipik örnek Laurus nobilis yapraklarında
bulunan eterik yağ hücreleridir.
197. • Şekil 31. Salgı hücreleri, A. Laurus (Defne) yaprağının enine
kesitinde eterik yağ hücresi; B. Ruta (Sedefotu) yaprağının
enine kesitinde kalsiyum oksalat kristali ihtiva eden hücre.
(Yakar, 1983’ den)
198. • 2. Süt boruları
Yumuşak ve selüloz çeperli hücrelerden
meydana gelirler. Salgılarına lateks adı verilir.
Lateksin bileşiminde karbonhidratlar,
glikozitler, alkaloidler, kauçuk vb. bulunur.
Meydana geliş şekillerine göre iki gruba
ayrılırlar.
199. • a) Eklemsiz Süt Boruları
– Meristematik kökenli tek hücreden meydana
gelmiş,çok nukleuslu ve çok fazla uzayabilen
borulardır. En fazla Moraceae ve Euphorbiaceae’de
bulunur.
• b) Eklemli Süt Boruları
– Üst üste ve yan yana gelmiş hücrelerin aralarında
bulunana enine çeperlerin erimesi ile meydana gelen
dallanmış ve ağsı yapı gösteren borulardır. Papaver
rhoeas, Chelidonium majus, Cichorium intybus ve
Taraxacum macrolepium ’da çok bulunur.
200. • Şekil 32. Salgı boruları. A. Euphorbia (Sütleğenotu)
kökünden alınmış boyuna kesitte segmentsiz süt borusu;
B, Taraxacum (Karahindiba) kökünden alınmış boyuna
kesitte segmentli süt borusu (Yakar, 1983’ den)
201. Hücre Dışı Salgılar
• Bunlar,üç ana grupta toplanabilir.
– a) Hücre arası Boşluklara Boşaltan salgılar
– Bunlar, salgı kanalları veya salgı cepleri şeklindedir.
Salgı kanalları,salgı maddeleri ile dolu olan hücre arası
boşlukların üst üste gelerek meydana getirdikleri
uzun kanallardır. Örneğin;Pinus sp.’deki reçine
kanalları
– Salgı cepleri ise toplu olarak bulunan birçok salgı
hücresinin salgılarını akıttıkları oval veya yuvarlak
boşluklardır. Rutaceae ve Umbelliferae’deki eterik yağ
salgıları salgı cepleri tarafından salgılanır.
202. • Şekil 33. Salgı cepleri. A, Hypericum perforatum
(Binbirdelikotu) yaprağından alınmış enine kesitte
şizogen eterik yağ cepi; B, Ruta (Sedefotu) yaprağından
alınmış enine kesitte lizigen eterik yağ cepi.
203. • b) Epidermis Hücreleri Tarafından Selüloz Çeper
ve Kutikula Arasına Boşaltılan Salgılar
• Bu tip salgılar uçucu ve aromatik eterik yağ
salgılayan salgı tüylerinde bulunur. Salgı tüyleri
çoğunlukla sap ve boş kısımlardan meydana gelir.
Hücre çeperi selülozdan yapılmıştır. Bunların
kutikulaları da incedir. Salgı maddesi tüyün baş
kısmındaki hücreler ile kutikula arasına
toplanmıştır. Kutikula oldukça şişkindir.
Compositae ve Labiatae familyalarında görülür.
204. • c)Doğrudan Doğruya Dışarı Doğru Boşaltan
Salgılar
• 1.Hidatodlar
• Çok nemli ortamlarda yaşayan bitkilerde
bulunan ve değişime uğramış
epidermis,trikom ve stomaya benzer
hücrelerden meydana gelmiş yapılardır.
Görevleri bitkilerde bulunan fazla suyu dışarı
atmaktır.
205. • Tranpirasyon azalıp su basıncı arttığı zaman su
hidatodlardan dışarı atılır. Pasif ve aktif hidatod olmak
üzere iki tip vardır. Pasifi hidatodlarda kök basıncı arttığı
zaman su,iletim sisteminin etkisiyle dışarı verilir. Hidatodun
kendisi salgılamaya katılmaz. Bunlardan stomaya benzeyen
açıklığın altında ince çeperli, hücre arası boşluklar
yönünden zengin, kloroplastsız, parenkimatik bir doku
bulunur. Bu dokuya epitema adı verilir. Bu tip hidatodlarda
su iletim borularının uçları epitema ile temas halindedir.
Pasif hidatodlar en fazla Primula sp. ve Fuchsia sp.’de
yaprak kenarlarında bulunur.
• Aktif hidatodlar metamorfoza uğramış tüylerdir. Suyun
dışarıya verilmesi tüyün kendi faaliyeti ile olmaktadır. İletim
sistemi ile doğrudan ilgileri yoktur. Örneğin. Phaseolus
vulgaris’in yaprak ve petiollerinde bulunan tüyler.
207. • 2. Enzim Bezleri
• Drosera sp., Dionea sp., Pinguicula sp., Nepenthes
sp. gibi insektivor bitkilerde proteinleri hidroliz
eden enzimler bulunmaktadır. Bu bezler, pepsin
özelliğinde bir enzim ile organik bir asit
salgılayarak böceklerin proteinli kısımlarını
peptonlara dönüştürürler. Bunların özel bir tipi
granülsüz ER’de depolanan mirosinaz enzimidir.
Bu enzim,hücre yaralanınca izotiyosiyanat
meydana getirir.
210. • 3. Tuz Salgı Bezleri
• Halofitlerdeki bezlerdir. Su buharlaşınca bu tip
bitkilerin yaprakları üzerinde kristal halinde
tuzlar birikir. Örneğin Suaeda sp.,Salicornia sp.
211. (Tuz salgı bezleri; Salt tolerant mechanisms in single-cell C4 species Bienertia sinuspersici and
Suaeda aralocaspica (Chenopodiaceae ); Plant Science 176: 616-626, 2009)
212. • 4. Nektaryumlar
• Böceklerle tozlaşan bitkilerde nektar denilen
bir sıvı salgılanır. Bileşiminde yüksek oranda
karbonhidrat,daha az oranda eterik yağlar,
organik asitler, protein ve alkoloidler bulunur.
Bitkinin yaşı arttıkça nektar veriminde azalma
olur. Bitkilerde bulunduğu yere göre iki gruba
ayrılır.
213. • a) Çiçek Nektaryumları
• Kaliks ve korollada,reseptakulum,filament ve ovaryum
septumlarında bulunurlar. Septumdakiler septal
nektaryum adını alıp sinkarp ovaryum septumlarının
tam birleşmesi sonucu buradaki epidermis
hücrelerinin bez özelliği kazanmasıyla meydana
gelirler. Özellikle Yucca sp.’de bol bulunurlar. Çiçek
kısımlarında yüzeysel olarak bulunan nektaryumlar ise
basit ve farkılılaşmamış olan bezlerdir. Epidermis
hücrelerinden yoğun plazmaları,sindirimi ve papilloz
şekilleri ile ayrılırlar. Perigonal (periantta bulunan),
toral (reseptakular) ,staminal, ovaryal ve stillar diye
beşe ayrılır.
215. • b) Ekstrafloral Nektaryumlar
• Çiçek dışındaki her yerde bulunurlar. En çok da
yaprak, stipül ve petiolde bulunurlar. Vicia
faba stipüllerinde, Impatiens sultanii yaprak
dişlerinde, Prunus spinosa yaprak dişlerinde ve
Euphorbia sp.’de yaprak kenarlarında bulunur.
217. • Nektar salınış tiplerine göre nektaryumlar şu şekilde
sınıflandırılırlar
Nektar kutikula bulundurmayan ince çeperli epidermal
hücreler arasından difüzyon ile salınır.
Epidermal kökenli tüylerden salınır. Tüyler tek ya da
çok hücreli olabilirler.
Geniş epidermal hücrelerin uzamasıyla oluşan ve
papiller adı verilen karbonhidrat içeren yapılardan salınır.
Stoma ile salınır.
Kutikulanın parçalanması ile musilaj yapısnda nektar
bulunur.
Nektar iletiminde ER çok önemlidir. Nektar floemden
plasmodesmatalar yardımıyla nektar parenkimasına gelir ve
yukarıdaki mekanizmalarla salınır.
218. • Osmoforlar
• Orchidaeceae ve Araliaceae gibi familyalarda sepal ve
petalin epidermisinde bulunan kokulu,uçucu maddelerdir.
Osmoforlar salgı maddesini vermeden önce salgı
hücrelerinin epidermisi sitoplazma yönünden,epidermis altı
doku ise nişasta yönünden zengindir. Sonra sitoplazma ve
nişasta azalır ve vakuoller büyür. Osmoforlar,genelde
terpen yapısındadır. En yaygın olanları da 1,8-sineol,metil
salisilat ve benzil asetatatır. Osmoforların salgısı akşam
üzeri 5 civarında salınmaya başlar,gece 3’de salınma
maksimum olur ve gündüz saat 5’de salgı en az düzeye iner.
Özellikle erkek arılarla tozlaşan Ophrys sp.’de yaygındır.
222. Emme Sistemi
• Bitki yapısına besin maddelerinin alınmasını
sağlayan sistemdir. Emilen maddeler
su,inorganik maddeler ve organik
maddelerdir.
223. • A.Suyun Yeraltı Organlar Tarafından Emilmesi
• 1.Kök ve Kök Tüyleri
• Su emilimi için geniş bir yüzeye sahip olmalıdırlar.
Bunun için yan kökler ve kök tüyleri oluşmuştur. Köklerin en
genç ve en yaşlı kısımları dışında kök emici tüyleri vardır.
Emici tüyler suyu toprak partiküllerinin absorbsiyon kuvveti
kendi emme kuvvetlerine eşit oluncaya kadar devamlı
emerler. Emilen su kök eipidermisi ve kök korteksi
yardımıyla endodermis ve ksileme geçer.
• 2)Rizoidler
• Bryohyta ve Pteridophyta gametofitlerinde suyu kılcal
kuvvetle emen ve taşıyan kök tüylerini andıran yapıdaki
rizoidler bulunmaktadır.
224. • B.Suyun Toprak Üstü organlar Tarafından Emilmesi
• 1.Yapraklar
• Bromeliaceae familyasında olduğu gibi epifit bitkilerde
yapraklar rozet şeklindedir ve dip kısımları suyu huni gibi
toplamaktadır. Bu bölgede kalkan tüyler bulunmaktadır. Tüyün alt
tarafı emme olayının gerçekleştiği kutikulasız kısımdır. Kurak
mevsimlerde tüyün sap hücreleri turgorlarını kaybederler. Bu
safhada tüy çukurluğun içine çekilir ve girintisini kapatır. Üst
kısımda kutikula olduğundan buradan da su kaybı olmaz.
• 2.Hava Kökleri
• Orchidaceae gibi familyalarda havada sarkan ve kök tüyü
olmayan kökler bulunur. Bu tip köklerde korteksin dış kısmı su
emen kılcal sistem halini alır. Çeperlerinde delikler ve belli
bölgelerde helezoni kalınlaşmalar vardır. Bu tabakaya velamen
radikulum denir.
225. • C.Organik Madde Emilmesi
• 1.Ototrof Bitkilerde
• Bu bitkiler, embriyo devresinde genellikle parazittir ve ana bitkiden organik
madde sağlarlar. Poaceae familyasında bulunan skutellum adlı yapı endosperme
enzimler göndererek buradan organik madde sağlarlar. Skutellum, endosperm ve
embriyo arasındaki sınır tabakasıdır.
• Monocotyledones tohumlarında da kotiledonlar endospermlerden besin
alırlar. Çimlenme devresi sona erince ise bitkilerde fotosentez başlar ve parazitlik
ortadan kalkar.
• 2.Heterotrof Bitkilerde
• Bunlarda organik madde emilimi hayat boyu devam eder. Peronospora sp.
ve Albugo sp. hifleriyle konak bitkinin boşluklarına yayılarak buralardan besin
sağlarlar.
• Hemiparazit olan Viscum album apoplastik süreklilik vasıtasıyla organik
madde sağlar. Holoparazit olan ve bitki gövdelerinde yaşayan Cuscuta sp. ‘de emeç
kökleri yardımıyla hem konağın ksilemine girerek su alır, hem de konak
floeminden anorganik ve organik madde sağlar.