Downloaded 204 times











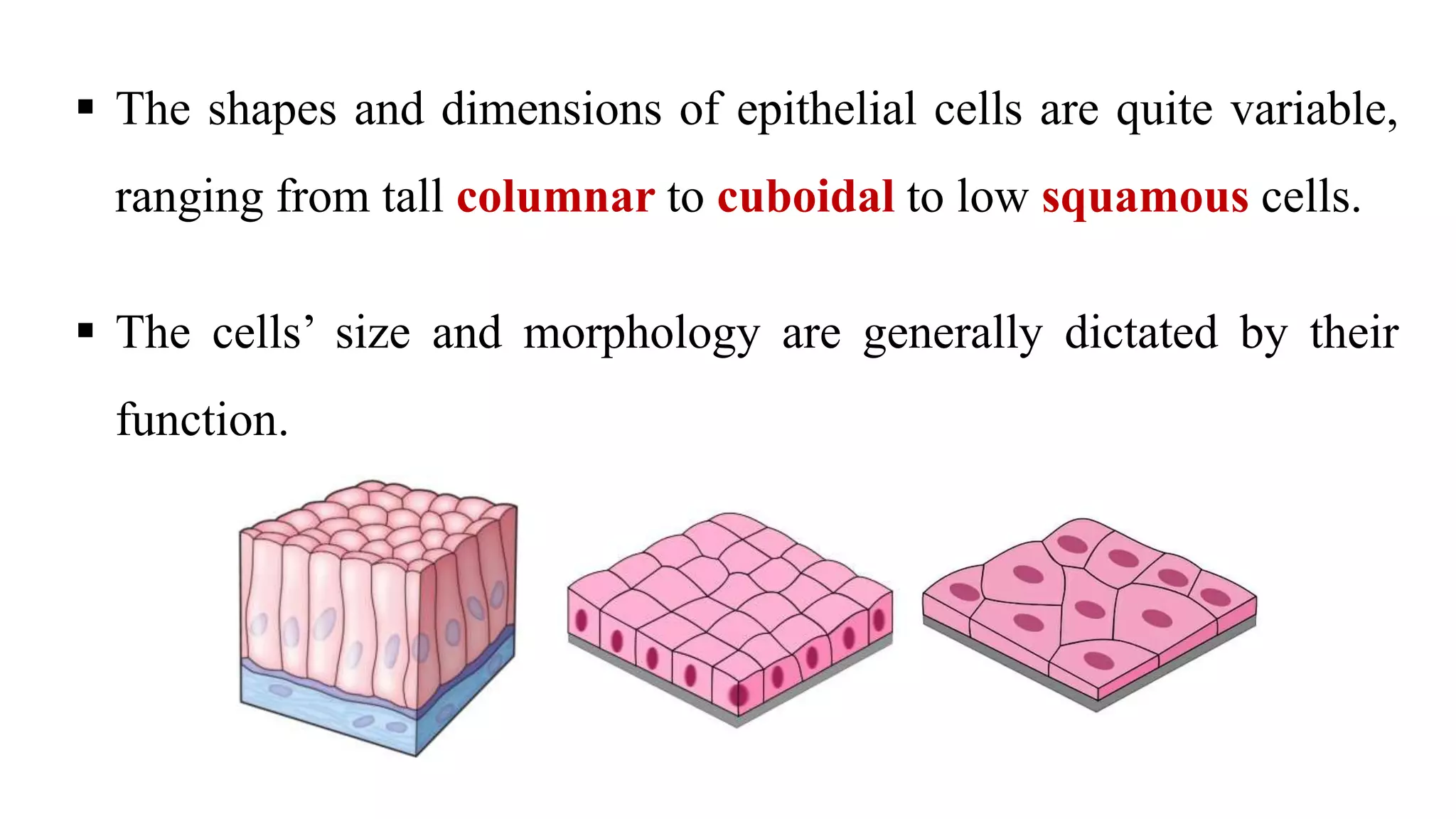




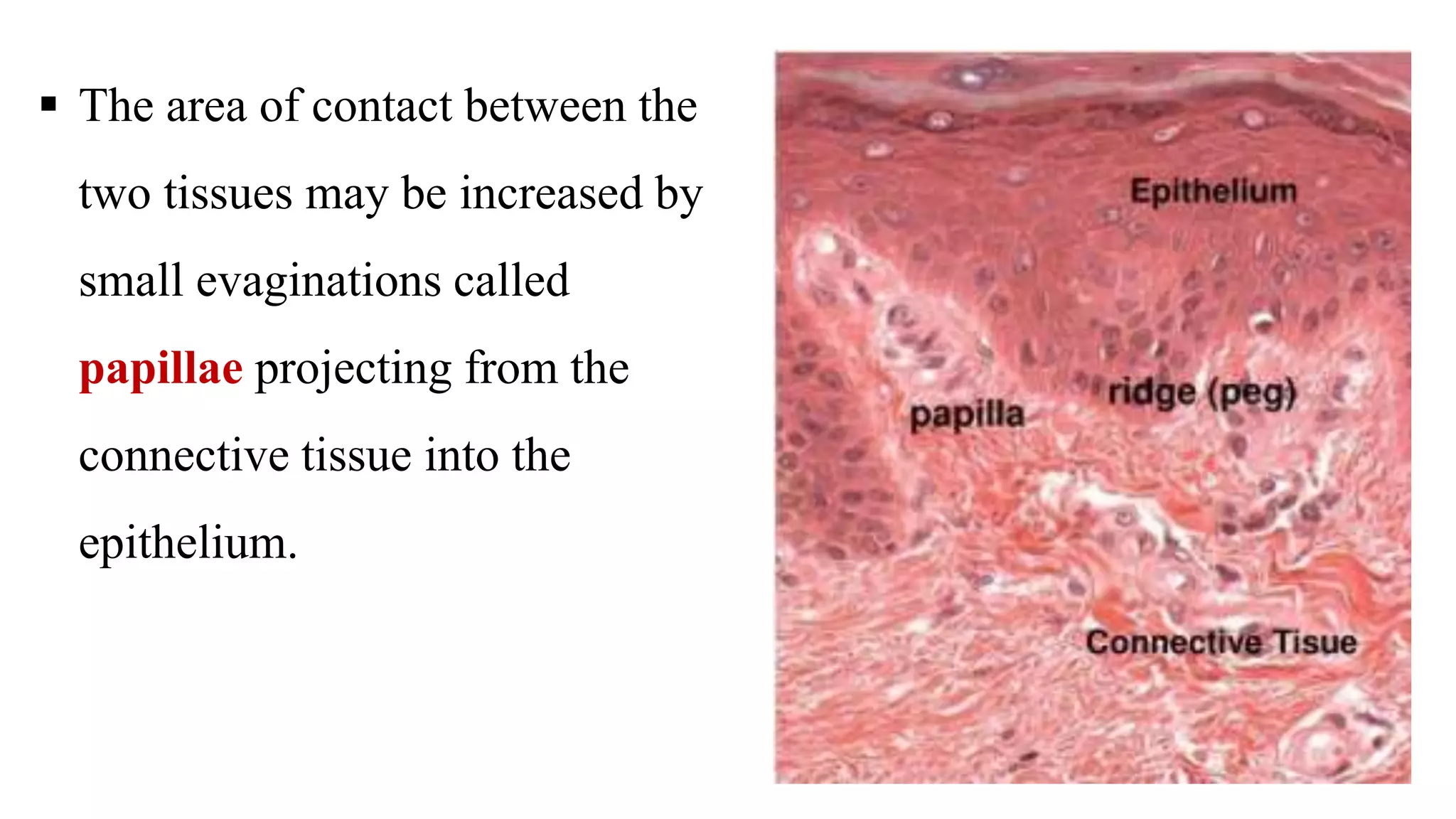












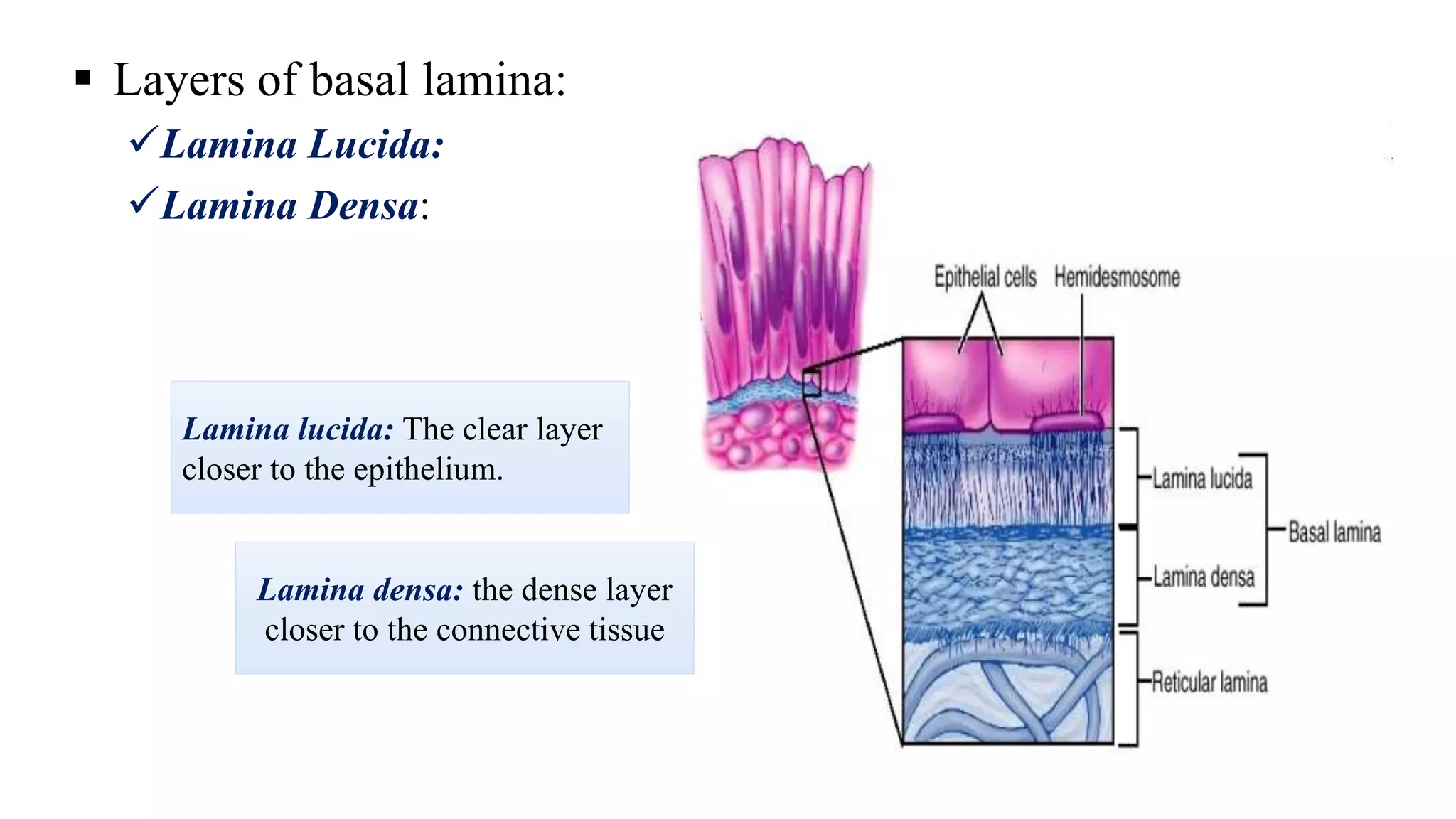










































































































































































Epithelial tissue is one of the four fundamental tissues in the human body, forming protective cellular layers that line external and internal surfaces, allowing for functions such as protection, secretion, absorption, and sensation. This document details the classification, structure, and functions of epithelial tissue, highlighting aspects such as intercellular junctions, the basement membrane, and specialized cell surfaces, including microvilli and cilia. Additionally, it discusses the medical significance of epithelial junctions and diseases related to their dysfunction.






























![Body_Tissues fully detailed notes [1].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/bodytissues1-250917032454-2d23aff7-thumbnail.jpg?width=640&height=640&fit=bounds)























































