Downloaded 170 times



![Michaelis and Menton Equation
• In MM expression
Total enzyme concentration= [ET] = [E] + [ES]
Free enzyme conc [E] = [ET] - [ES]
Substrate concentration = [S]
Initial velocity = Vo, Velocity measured immediately after
mixing E + S, at beginning of reaction (initial velocity), is
called Vo.
Maximum velocity = Vmax
Half Vmax = Km (substrate concentration)
Km = substrate concentration that gives Vo = 1/2 Vmax.](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-4-2048.jpg)
![At very low [S]:
• Vo is proportional to [S];
doubling [S] → double Vo.
2. In mid-range of [S], Vo is
increasing less as [S]
increases (where Vo is
around 1/2 Vmax).
Km = [S] that gives Vo = 1/2
Vmax.
3. At very high [S], Vo is
independent of [S]:
Vo = Vmax.
Enzyme-catalyzed reactions show a hyperbolic
dependence of Vo on [S]](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-5-2048.jpg)
![Derivation
• Initial velocity Vo= k2[ES]
• Rate of formation of [ES] = k1 [E][S]
= k1([ET]-[ES]).[S]
= k1[ET][S] - [ES][S]
• Rate of breakdown of [ES] =k-1[ES] + k2[ES]
= (k-1+k2)[ES]
• Steady state:
Rate of formation = Rate of breakdown
k1[ET][S] - [ES][S] = (k-1+k2)[ES]](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-6-2048.jpg)
![• separation of rate constants
k1[ET][S] - [ES][S] = (k-1+k2)[ES]
k1[ET][S] = [ES][S] + (k-1+k2)[ES]
k1[ET][S] = {[S] + (k-1+k2)} [ES]
[ES] = k1[ET][S]
[S] + (k-1+k2)
[ES] = [ET][S]
[S] + k-1+k2
k1
k-1+k2 = Km [ES] = [ET][S] ......... (i)
k1 [S] + Km](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-7-2048.jpg)
![• Expressing Vo in term of [ES]:
multiply k2 on both side of eq (i)
k2. [ES] = k2 [ET][S] ...... (ii)
[S] + Km
As we know Vo= k2[ES]
So, eq (ii) becomes
Vo = k2 [ET][S] .....(iii)
[S] + Km
When [S] is greater, then Vo becomes Vmax and
Vmax= k2 [ET]
So, eq (iii) becomes
Vo = Vmax [S]
[S] + Km](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-8-2048.jpg)
![Michaelis-Menten equation
explains hyperbolic Vo vs. [S]
curve:
1. At very low [S] ([S] << Km), Vo
approaches (Vmax/Km)[S]. Vmax and
Km are constants, so linear
relationship between Vo and [S]
at low [S].
2. When [S] = Km, Vo = 1/2 Vmax
3. At very high [S], ([S] >> Km), Vo
approaches Vmax (velocity
independent of [S])
Explaination of hyperbolic curve](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-9-2048.jpg)
![Significance of MM equation
It describes
• kinetic behaviour of enzymes
• hyperbolic dependence of Vo on [S]
• independance of number of steps involved
• different enzymes have different Km and Vmax.
• Km and Vmaxmay be influenced by pH, temperature.
• Km can be used as a relative measure of the affinity of the
enzyme for each substrate (smaller Km means higher
affinity)
• in a metabolic pathways, Km values may indicate the rate-
limiting step (highest Km means slowest step).
• Vmax is independent of [S] at saturation.](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-10-2048.jpg)

![Lineweaver-Burk plot
• A more convenient graphical representation of MM
equation
• It is a straight line plot, easier to evaluate than curves.
• Lineweaver-Burk plot is a double reciprocal plot obtained
by taking reciprocal of both sides of MM equation and
rearranging
1 = Km + [S]
V [S] Vmax
1 = Km 1 + 1
V Vmax [S] Vmax](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-12-2048.jpg)
![• A plot of 1/V versus 1/[S] is a straight line having a slope
of Kmax/Vmaxand an intercept of 1/Vmax on the y-axis](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-13-2048.jpg)

![Reversible Inhibition
• It is further subdivided into competitive, noncompetitive
and uncompetitive types.
• In reversible inhibition, equilibrium exists between
inhibitor I and enzyme E as
E+I EI
• The eq. constant for the dissociation of EI complex,
called Ki is given by equation
Ki = [E][I]
[EI]
• Ki is the measure of affinity of the inhibitor for enzyme
simliar to Km.](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-15-2048.jpg)
![Competitive inhibition
• The inhibitor is a structural analogue that competes with
the substrate for binding at active site.
• Because the inhibitor binds reversibly to the enzyme,so
when [S] far exceeds [I], the probability that an inhibitor
molecule will bind to the enzyme is minimized and the
reaction exhibits a normal Vmax.](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-16-2048.jpg)
![Noncompetitive inhibition
• Inhibitor does not usually bear any structural
resemblance to the subatrate and it binds to the enzyme
at a site distinct from the substrate binding site.
• No competition exists between inhibitor and substrate,
so inhibition cannot be overcome by increase of [S].
• Vmax is reduced by inhibitor but Km is unaffected
because the affinity of S for E is unchanged.](https://image.slidesharecdn.com/enzymekinetics-171022075327/75/Enzyme-kinetics-17-2048.jpg)
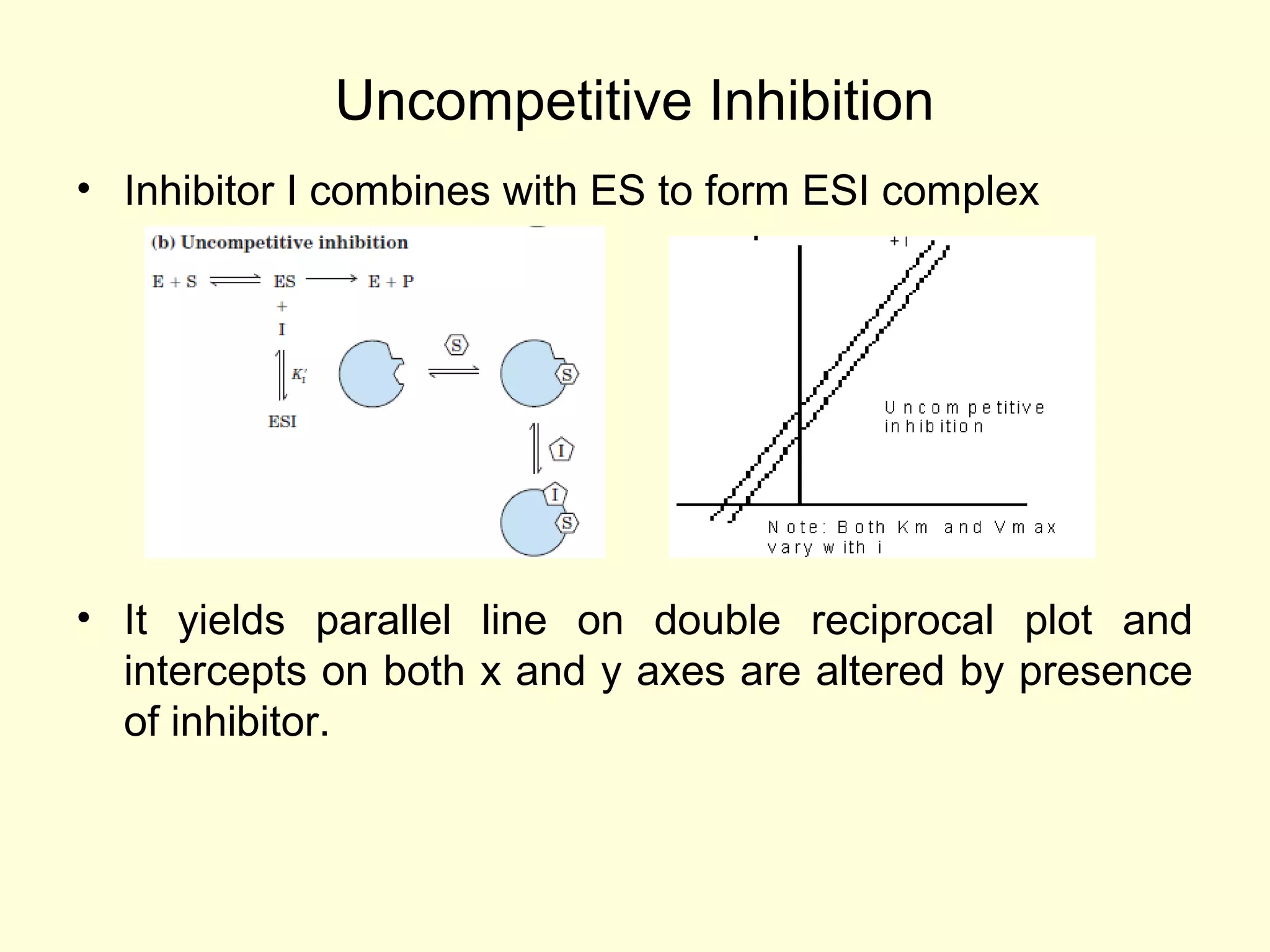

Enzyme kinetics is the study of enzyme-catalyzed reaction rates. The Michaelis-Menten equation relates reaction velocity to substrate concentration and kinetic parameters. It describes the hyperbolic relationship between velocity and substrate concentration. The equation can be linearized into the Lineweaver-Burk plot for easier analysis. Enzyme inhibition studies help understand reaction mechanisms and are important for drug development, as most drugs function by inhibiting specific enzymes.























































































