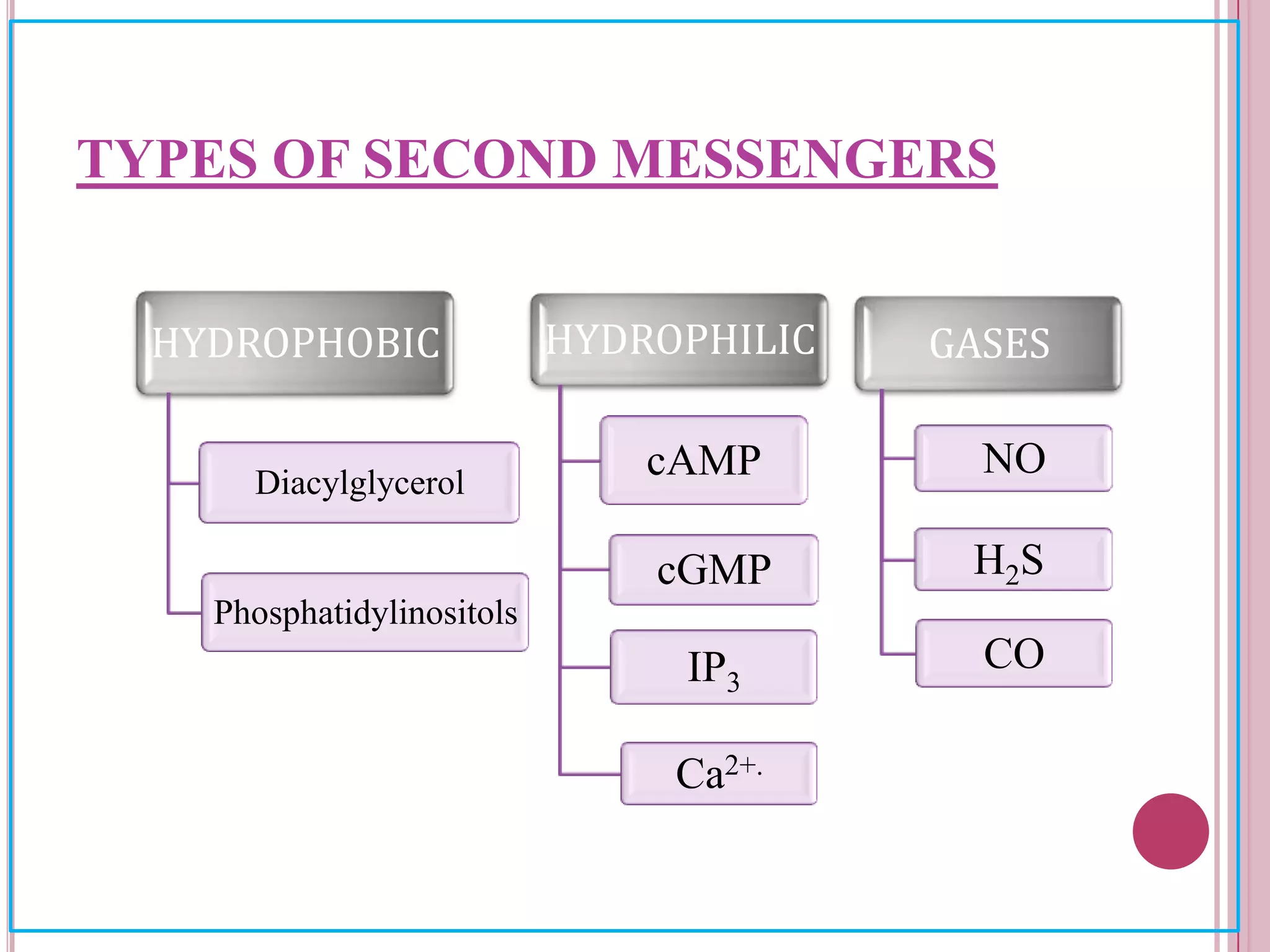
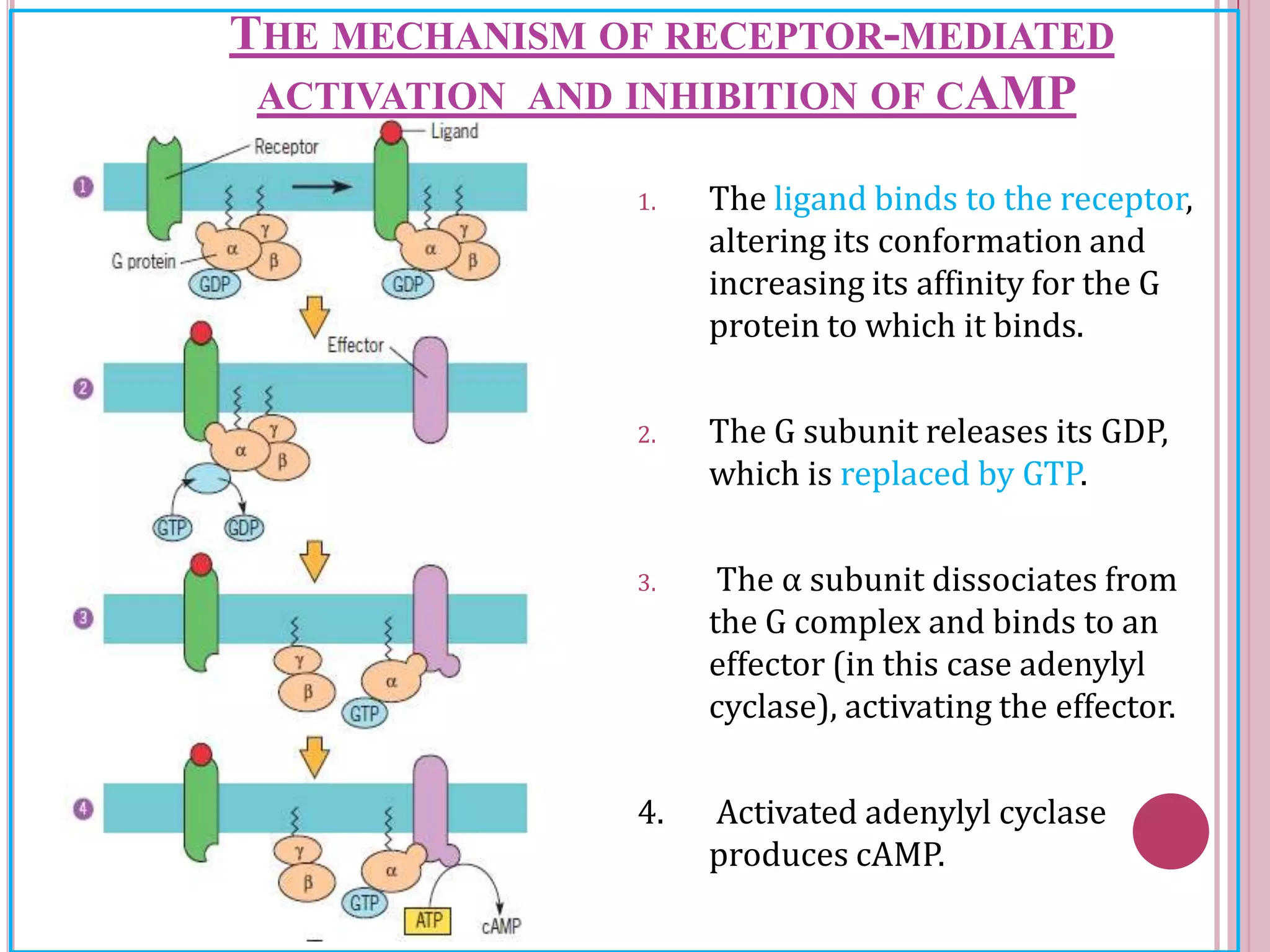
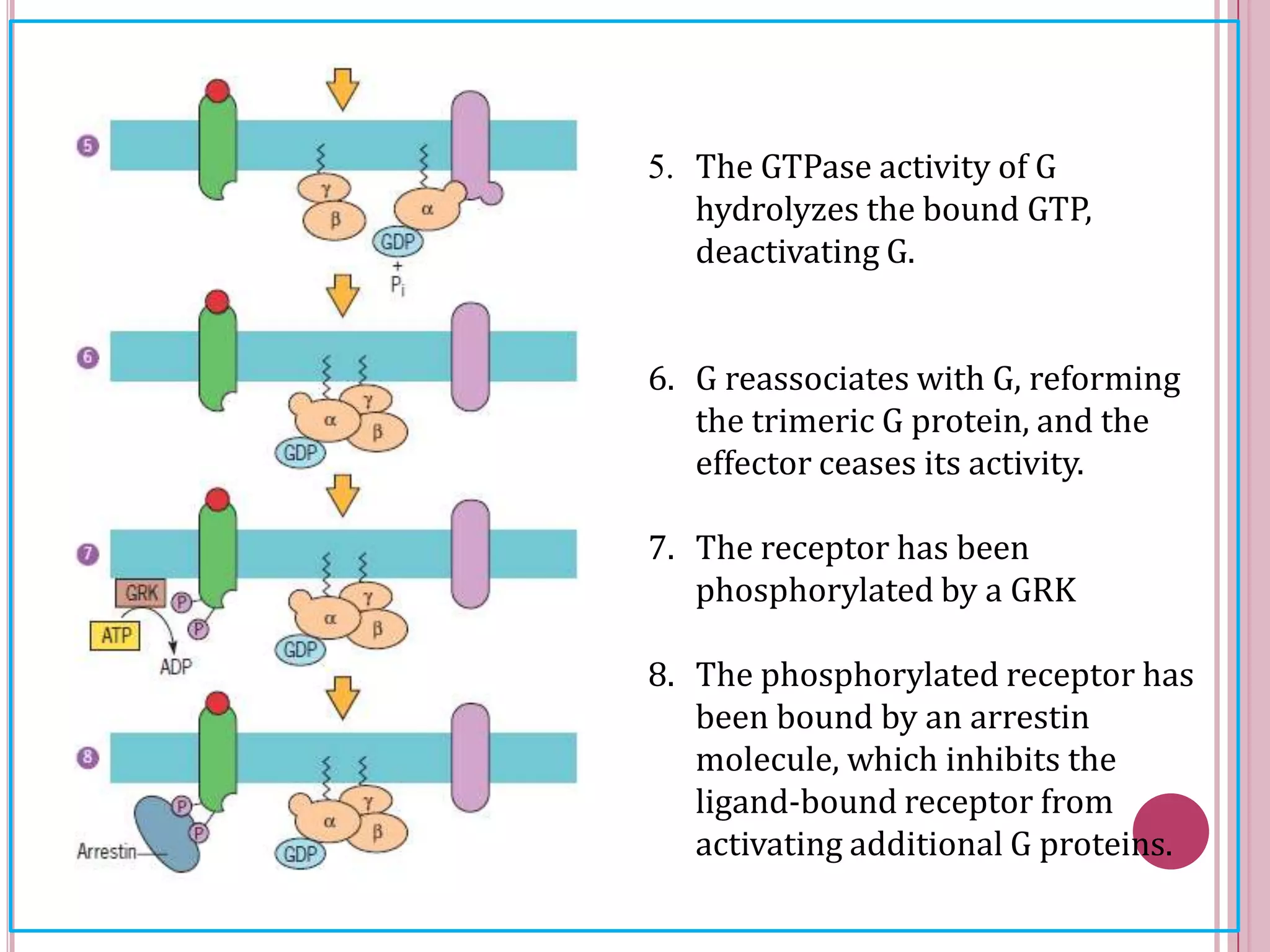
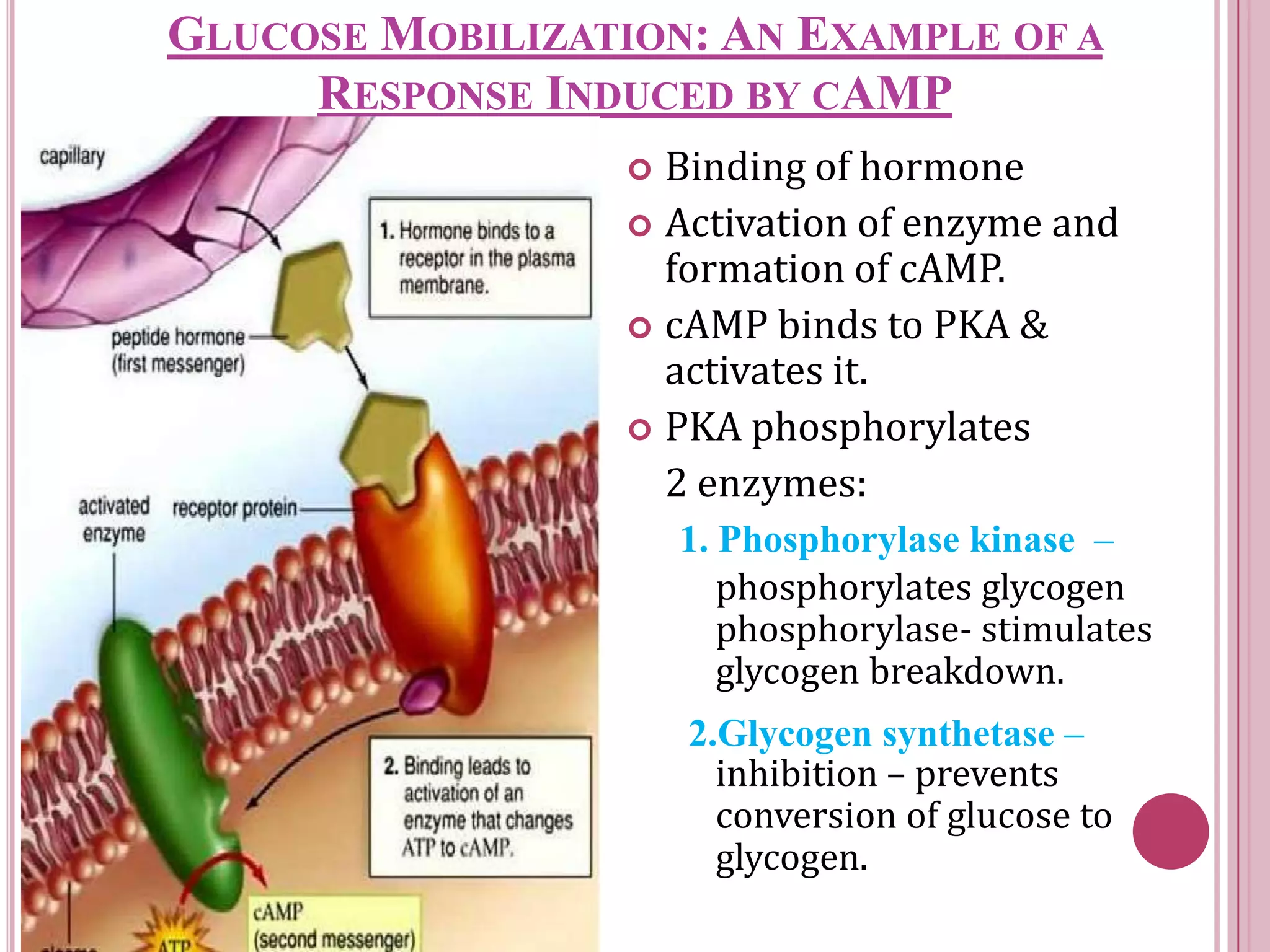
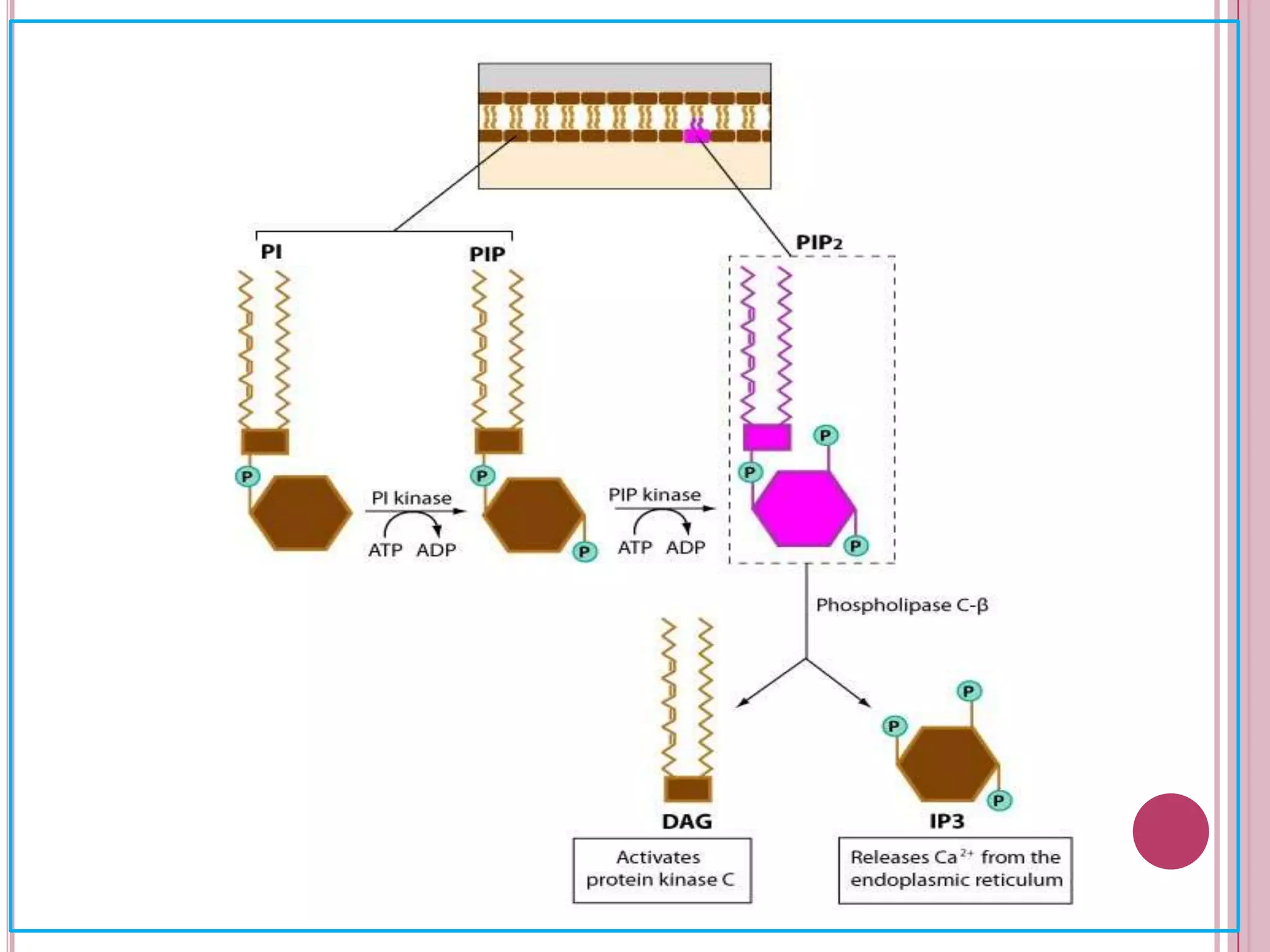
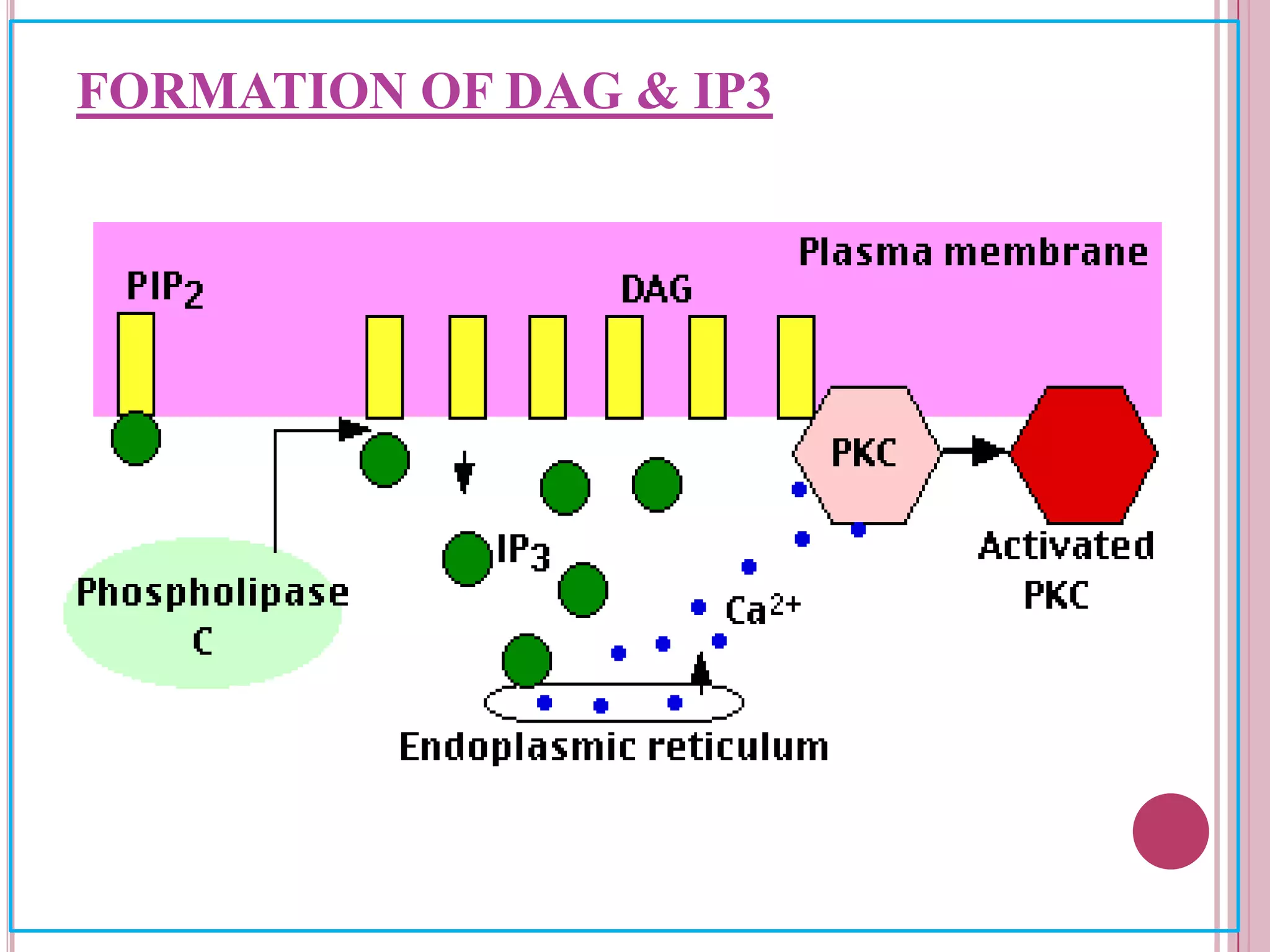
Second messengers are small intracellular molecules that amplify signals received at cell surface receptors and help transmit them to target molecules inside the cell. The document discusses four main classes of second messengers - cyclic nucleotides, membrane lipid derivatives, calcium ions, and gases like nitric oxide. It provides details on several important second messengers, including cAMP, cGMP, IP3, DAG, and calcium ions, and how they mediate intracellular signaling pathways and cellular responses.