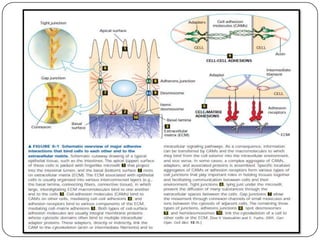
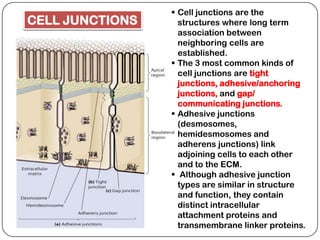
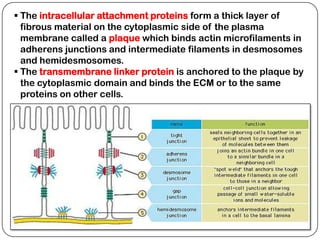
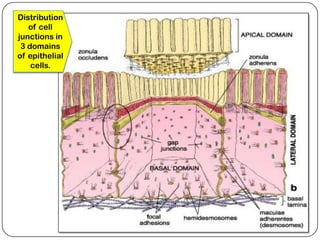
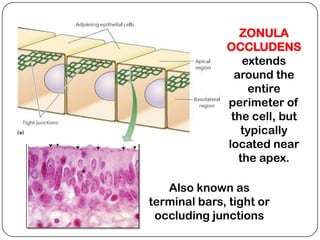
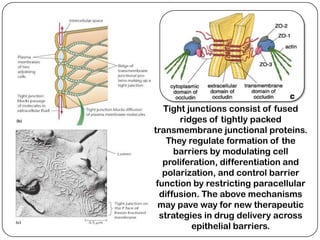
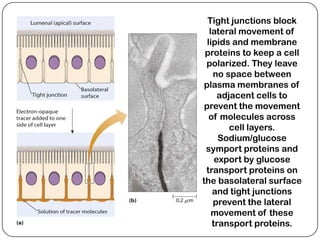
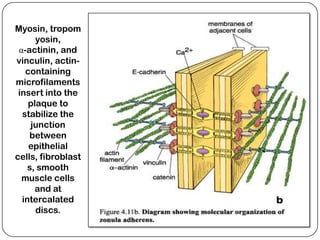
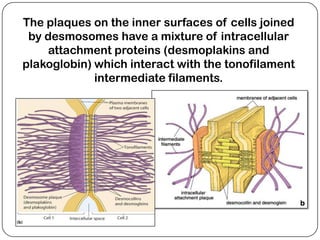
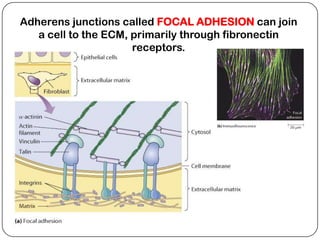
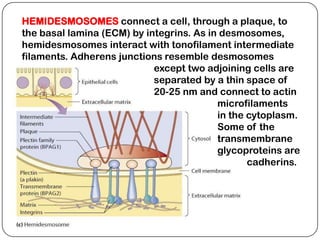
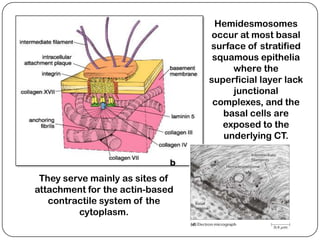
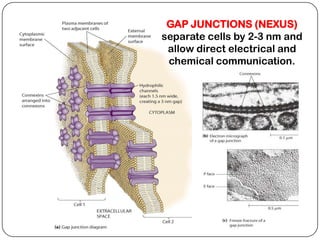
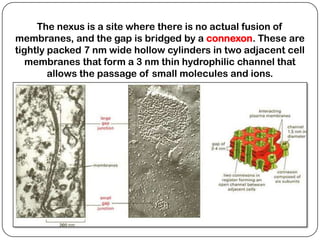
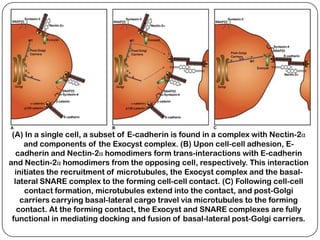
Cell junctions are structures that allow neighboring cells to associate with each other. The three main types of cell junctions are tight junctions, adhesive junctions, and gap junctions. Adhesive junctions like desmosomes and adherens junctions link cells together and to the extracellular matrix. These junctions contain intracellular attachment proteins and transmembrane linker proteins that anchor the cells. Gap junctions allow direct communication between cells by forming channels that let small molecules pass between cells. Cell junctions play important roles in cell polarization, barrier function, and coordinated cell behavior.