Sarcomero
•Download as PPTX, PDF•
0 likes•172 views

El sarcómero es la unidad funcional de la musculatura esquelética.

Recommended
More Related Content
What's hot
What's hot (20)
Similar to Sarcomero
Similar to Sarcomero (20)
More from ZORAIDA BEATRÍZ FERNÁNDEZ MARTÍN
More from ZORAIDA BEATRÍZ FERNÁNDEZ MARTÍN (10)
Recently uploaded
Recently uploaded (20)
Sarcomero
- 1. Dra. Zoraida Beatríz Fernández Martín SARCÓMERO
- 2. SARCÓMERO Las miofibrillas son haces de filamentos gruesos y delgados organizados en unidades funcionales repetitivas llamadas sarcómeros y como en reposo cada uno de ellos tiene una longitud de 1.6-2.6 µm, una miofibrilla tiene aproximadamente 10,000 sarcómeros de un extremo a otro. Los sarcómeros son las unidades funcionales mas pequeñas de la fibra muscular y cada uno de ellos contiene filamentos gruesos, filamentos delgados, proteinas que estabilizan la posición de los filamentos y proteinas que regulan las interacciones entre los filamentos delgados y gruesos. Las interacciones entre los filamentos gruesos y delgados de los sarcómeros son las responsables de la contracción muscular.
- 3. SARCÓMERO La apariencia estriada de cada miofibrilla se debe a las diferencias en tamaño, densidad y distribución de los filamentos gruesos y delgados. La estriación se forma alternando bandas oscuras (bandas-A) y claras (bandas-I), cuyos nombres derivan de las palabras anisotrópico e isotrópico y se refieren a la apariencia de las bandas cuando son vistas con un microscopio de luz polarizada.
- 4. MIOFIBRILLAS Los componentes primarios de las fibrillas de músculo esquelético son miosina y actina, así como tropomiosina y troponina asociadas con la actina. Otras proteinas musculares, como titina, neblina, α-actina y miomesina, son esenciales para regular el espaciamiento, unión y alineación precisa de los miofilamentos. Estas proteinas constituyen mas del 75% del total de proteinas de la fibra muscular.
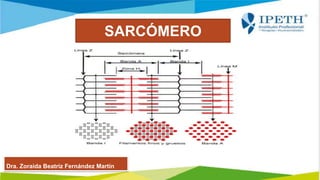
- 6. La banda A. Los filamentos gruesos están colocados en el centro del sarcómero, en la banda-A, cuya longitud es igual a la longitud de un filamento grueso típico. Sin embargo, la banda-A también incluye porciones de los filamentos delgados y tiene tres subdivisiones: La línea-M. La porción central de cada filamento grueso se conecta con sus vecinos por medio de proteínas en la línea-M, que ayudan a estabilizar sus posiciones. La zona-H. En un Sarcómero en reposo, la zona-H o banda-H es una región mas clara a cada lado de la línea-M y contiene filamentos gruesos pero no filamentos delgados La zona de sobreposición. En esta zona los filamentos delgados están situados entre los filamentos gruesos, de manera que cada filamento delgado está rodeado por tres filamentos gruesos y cada filamento grueso por seis filamentos delgados. LA BANDA- A
- 7. LA BANDA- I • Cada banda-I se extiende desde la banda-A de un Sarcómero hasta la banda-A del siguiente sarcómero y contiene filamentos delgados, pero no filamentos gruesos, con las líneas-Z marcando la frontera entre sarcómeros adyacentes. • Estas líneas-Z consisten de proteínas llamadas 'conectinas', que interconectan los filamentos delgados de sarcómeros adyacentes. • Desde las líneas-Z en el extremo de cada Sarcómero, los filamentos delgados se extienden en la zona de sobre posición hacia la línea-M. Hilos de una proteina llamada 'titina' se extienden desde la punta de los filamentos gruesos a los sitios de unión en la línea-Z y ayudan a la fibra muscular a resistir el estiramiento, que en otra forma trastornaría el arreglo de miofibrillas y el mecanismo de la contracción. • Cada Sarcómero está rodeado por dos túbulos transversos y las triadas se localizan a ambos lados de la línea-M, en la zona de sobre posición. Como resultado, los iones calcio liberados por el retículo sarcoplásmico entran en las regiones donde los filamentos delgados y gruesos interactúan. • Cada línea-Z está rodeada por una red de filamentos intermedios que interconectan miofibrillas adyacentes y las mas cercanas al sarcolema se unen a sitios específicos en el interior de la membrana. Debido a que las líneas-Z de todas las miofibrillas están alineadas, la
- 8. SARCOLEMA El sarcolema o membrana celular rodea el sarcoplasma o citoplasma de la fibra muscular y como otras membranas celulares, puede mantener un potencial a través de ella debido a la diferencia en la concentración de cargas positivas y negativas en el interior y exterior de la célula. El primer paso en el proceso que lleva a la contracción es un cambio brusco en el potencial transmembrana, llamado potencial de acción.
- 9. SARCOLEMA Tiene numerosas caveolas, consideradas como un tipo especializado de regiones lipídicas llamadas 'balsas', que contienen la proteína caveolina-3, específica del músculo. En el músculo esquelético diferenciado la caveolina- 3 se asocia con los túbulos-T en desarrollo, pero en el músculo maduro está restringida a las caveolas del sarcolema y ya no es detectable en las túbulos-T La localización exacta de caveolina-3 respecto a marcadores definidos de superficie ha sido estudiada por Rahkila y colaboradores (2001), quienes encontraron que el sarcolema del músculo parece ser un arreglo de áreas con balsas asociadas con caveolina y áreas sin balsas, que forman un mosaico de dominios de túbulos-T, vacuolas sarcolemales y dominios de β- distroglicanos.
- 10. RETÍCULO SARCOPLASMICO Donde un túbulo transverso encuentra una miofibrilla, el túbulo es rodeado de cerca por la membrana del retículo sarcoplásmico (RS). Este es un complejo membranal similar al retículo endoplásmico en otras células, pero en el músculo esquelético forma una red tubular alrededor de cada miofibrilla. A cada lado del túbulo-T el retículo sarcoplásmico se ensancha y forma una cámara llamada cisterna terminal, que se une al túbulo-T por medio de una estructura conocida como 'pie'. La combinación de un par de cisternas terminales y el tubulo transverso se denomina 'triada' y aunque sus membranas están unidas, los contenidos líquidos están separados y son diferentes.
- 11. RETÍCULO SARCOPLASMICO La concentración intracelular de iones calcio (Ca2+) se mantiene baja debido a 'bombas', que los sacan cuando su concentración aumenta. Aunque las fibras de músculo esquelético 'bombean' Ca2+ hacia afuera de la célula, también eliminan el calcio del sarcoplasma transportándolo a la cisterna terminal del retículo sarcoplásmico. El sarcoplasma de una fibra muscular en reposo contiene concentraciones de Ca2+ alrededor de 10-7 meq/litro y su concentración dentro de la cisterna terminal puede ser hasta 1000 veces mayor; además, la cisterna contiene una proteína llamada calsecuestrina, que une los iones calcio en forma reversible. Incluyendo tanto el calcio libre como el calcio unido a otras moléculas, la concentración total de Ca2+ dentro de la cisterna puede ser hasta 40,000 veces la del interior del sarcoplasma que la rodea.
- 12. RETÍCULO SARCOPLASMICO Para que ocurra la interacción entre la actina y la miosina que produce la contracción, debe haber calcio, que después de la contracción debe ser eliminado y la entrega y eliminación de este ión se lleva a cabo por el trabajo combinado del sistema tubular-T y el RS. El RS rodea las miofibrillas como un sistema de redes, una de ellas alrededor de la banda-A y otra en la banda-I, y donde las dos redes se encuentran, en la unión de las bandas-A e -I, el RS forma una cisterna. El RS controla el nivel de Ca2+ intracelular en el músculo esquelético, almacenándolo y liberándolo.
- 13. RETÍCULO SARCOPLASMICO Inicialmente el retículo sarcoplásmico se desarrolla como un retículo endoplásmico, pero conforme el músculo se diferencia, se enriquece con proteínas específicas. Tres proteínas purificadas inicialmente del retículo sarcoplásmico son: - ATPasa de calcio (SERCA. Es responsable de bombear calcio hacia la luz del RS durante la relajación. La mas abundante fuera de la unión túbulo-T-RS es SERCA, que normalmente está distribuida en elementos tubulares rodeando la línea-Z y -M, así como en los elementos alineados con el eje longitudinal de la miofibrilla. - Calsecuestrina (CLQ. Es la mas prominente de las proteínas intraluminales quelantes de calcio y entre ambas aumentan la capacidad del RS para el calcio. En la luz del RS la proteína mas abundante es CLQ. - Receptor a la rianodina (RyR). Es proteína ácida que une el calcio con afinidad moderada y alta capacidad.
- 14. SISTEMA TUBULAR TRANSVERSO (STT) Los túbulos-T forman una red continua que corre transversalmente por varias miofibrillas y las penetra a todos los niveles, siendo sistemas de membranas conectadas con el sarcolema que mantienen una composición de lípidos y proteínas diferentes a la del sarcolema Una fibra muscular esquelética puede ser muy larga, hasta de varios centímetros de longitud, pero todas sus regiones deben contraerse simultáneamente, por lo que la señal que inicia la contracción debe distribuirse rápidamente a lo largo y hacia el interior de la célula. Para ello, la señal es propagada primero a lo largo del sarcolema y después conducida por los túbulos transversos o túbulos-T, Son tubos estrechos continuos con el sarcolema que se extienden en ángulos rectos a la superficie celular. Los túbulos-T están llenos de líquido extracelular y forman vías dentro de la fibra muscular. Como su membrana tiene las mismas propiedades generales que el sarcolema, los potenciales de acción son conducidos hasta llegar a la región de las triadas, donde se inicia el proceso que acopla la excitación con la contracción. Aunque esta función de propagar rápidamente los potenciales de acción en la membrana superficial es la mejor conocida. Los túbulos-T también pueden ser utilizados para llevar componentes del líquido extracelular al interior de las fibras musculares.
- 15. TÚBULOS T • Los túbulos-T están localizados entre la cisterna terminal adyacente al RS, formando una triada compuesta por dos cisternas terminales y el túbulo-T. Las uniones RS-túbulo- T y sus asociaciones con miofibrillas se desarrollan en una serie de pasos consecutivos, donde la formación de uniones entre los dos sistemas membranales se hace en forma concurrente, iniciando cambios moleculares en ambos sistemas de membranas
- 16. TÚBULOS T Cuando el potencial de acción nervioso llega a la membrana muscular, esta sufre una despolarización que es trasmitida a las profundidades de la célula por las membranas del sistema tubular-T. Cuando el potencial de acción de los túbulos es detectado por un receptor a la dihidropiridina localizado en la unión del túbulo con la cisterna del retículo sarcoplásmico, un mecanismo aún no dilucidado produce la apertura de los receptores a la rianodina, que funcionan como canales para el calcio. Cuando estos canales se abren, como la concentración de iones calcio dentro del RS es mucho mayor que en citosol de la célula, estos iones salen de su depósito para iniciar el proceso de la interacción entre los filamentos gruesos y delgados, la contracción muscula Esta serie de eventos es llamada el 'acoplamiento excitación-contracción' (e-c) y varias proteínas específicamente localizadas en la unión RS-túbulo-T tienen papeles esenciales en ellos. El receptor a la dihidropiridina (DHPR) en el túbulo-T detecta el voltaje a través de la membrana y su activación produce la liberación de Ca2+ del RS (Flucher, 1992), mientras el canal de liberación RyR/Ca2+ está localizado en el RS de la unión y es responsable de la liberación de Ca2+ de sus lugares de almacenamiento. Tanto el RyR como el DHPR son necesarios para el desarrollado apropiado del músculo, aunque ninguno de ellos lo es para el anclaje T-RS o la búsqueda y/o asociación de CLQ y tradina en el RS de unión (Felder y Franzini-Armstrong, 2002).