ACIDOS NUCLÉICOS 2º Bachillerato - Ver.2014
•Descargar como PPT, PDF•
0 recomendaciones•1,719 vistas

ACIDOS NUCLÉICOS

Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Destacado (20)
Similar a ACIDOS NUCLÉICOS 2º Bachillerato - Ver.2014
Similar a ACIDOS NUCLÉICOS 2º Bachillerato - Ver.2014 (20)
Más de Javier García Calleja
Más de Javier García Calleja (18)
Último
Último (20)
ACIDOS NUCLÉICOS 2º Bachillerato - Ver.2014
- 5. Nucleótido.
- 9. Nucleótidos que son coenzimas. ATP NAD NADP FAD Coenzima A
- 10. ATP.
- 11. ATP, ADP, AMP.
- 12. ATP y obtención de energía. Se puede representar así: A-P~P~P Donde “~” son los enlaces anhídrido de ácido, que son de alta energía. En la hidrólisis del ATP se está hidrolizando uno de esos enlaces anhídrido de ácido. Esto libera gran energía, concretamente 7'7kcal/mol.
- 13. Polinucleótidos.
- 15. Los descubridores. En los años 20, el bioquímico Phoebus Levene determinó que el DNA estaba formado por 4 tipos distintos de nucleótidos. En 1949, el bioquímico Erwin Chargaff analizó el contenido molar de las bases de DNA procedente de diversos organismos y descubrió que en todos los casos [A]=[T] y que [G]=[C], o lo que es lo mismo, [A+G]=[T+C] ([purinas]=[pirimidinas]). Esta es la llamada ley de Chargaff. .
- 16. Los descubridores (II). A primeros de los años 50 Maurice Wilkins y Rosalind Franklin realizaron los primeros estudios físicos con el DNA mediante la técnica de difracción de rayos X y observaron que (1) la molécula de DNA es una cadena extendida con una estructura altamente ordenada (2) la molécula de DNA es helicoidal y tiene 20 Å de diámetro (3) la hélice del DNA está compuesta por dos hebras helicoidales y (4) las bases de los nucleótidos están apiladas con los planos separados por una distancia de 3,4 Å. Maurice Wilkins Rosalind Franklin Patrón de difracción de rayos X del DNA
- 17. ESTUDIOS DE DIFRACCIÓN DEL ADN.
- 18. Los descubridores (III). En 1953, James Watson y Francis Crick combinaron los datos químicos y físicos del DNA, y propusieron un modelo estructural del DNA que publicaron en la revista Nature. Watson y Crick Construyendo el modelo Construyendo el modelo
- 19. Un artículo indispensable (de sólo 2 páginas).
- 20. Según Watson y Crick... En este modelo estructural del DNA: las dos hebras están enrolladas una alrededor de la otra formando una doble hebra helicoidal : las dos cadenas de polinucleótidos se mantienen equidistantes, al tiempo que se enrollan en torno a un eje imaginario. El esqueleto azúcar-fosfato (formado por una secuencia alternante de desoxirribosa y fosfato, unidos por enlaces fosfodiéster 5'-3') sigue una trayectoria helicoidal en la parte exterior de la molécula. Las bases se dirigen desde cada cadena al eje central imaginario. Las bases de una hebra están enfrentadas con las de la otra, formando los llamados pares de bases (PB). Las bases interaccionan entre sí mediante puentes de hidrógeno. Las dos bases que forman un PB están en el mismo plano y dicho plano es perpendicular al eje de la hélice.
- 21. Según Watson y Crick... Los pares de bases están formados siempre por una purina y una pirimidina, de forma que ambas cadenas están siempre equidistantes, a unos 11 Å una de la otra. Los PB adoptan una disposición helicoidal en el núcleo central de la molécula, ya que presentan una rotación de 36º con respecto al par adyacente, de forma que hay 10 PB por cada vuelta de la hélice. La A se empareja siempre con la T mediante dos puentes de hidrógeno, mientras que la C se empareja siempre con la G por medio de 3 puentes de hidrógeno.El apareamiento de bases es una de las características más importantes de la estructura del DNA porque significa que las secuencias de bases de ambas hebras son complementarias. Este hecho tiene implicaciones muy profundas con respecto al mecanismo de replicación del DNA, porque de esta forma la réplica de cada una de las hebras obtiene la secuencia de bases de la hebra complementaria.
- 22. Según Watson y Crick... En cada extremo de una doble hélice lineal de DNA, el extremo 3'-OH de una de las hebras es adyacente al extremo 5'-P (fosfato) de la otra. En otras palabras, las dos hebras son antiparalelas, es decir, tienen una orientación diferente. Por convención, la secuencia de bases de una hebra sencilla se escribe con el extremo 5'-P a la izquierda. La doble hélice es dextrógira. Esto quiere decir que si alguien mira al eje de la hélice hacia abajo, en cualquier dirección, cada una de las hebras sigue una trayectoria en el sentido de las agujas del reloj al alejarse del observador. La hélice presenta dos tipos de surcos helicoidales externos, unos son anchos y profundos (surcos mayores) y otros son estrechos y poco profundos (surcos menores). Los dos tipos de surcos son lo suficientemente amplios como para permitir que las moléculas proteicas entren en contacto con las bases.
- 23. Según Watson y Crick... La relación A=T y C=G siempre se cumple, pero no hay regla que rija las concentraciones totales de G+C y de A+T. Existe una enorme variación en esta relación para los diferentes tipos de bacterias. Normalmente la composición de bases de una molécula de DNA de un organismo se expresa como su contenido en G+C. En organismos superiores, este valor está próximo a 0,5, pero en los organismos inferiores (bacterias) este valor varía mucho de un género a otro. Así, en el género Clostridium es de 0,27; en Sarcina es de 0,76 y en Escherichia es de 0,5. Esta estructura de doble hélice del DNA no es la más habitual. En las células eucariotas, el DNA se encuentra localiado principalmente en el núcleo, en forma de cromosomas, que son complejas asociaciones de DNA y proteínas.
- 24. Polinucleótidos.
- 25. En el ADN las bases son complementarias.
- 26. Y dos cadenas antiparalelas se unen entre sí.
- 27. ADN.
- 28. ADN.
- 29. ADN.
- 30. FORMAS DEL ADN.
- 31. Nucleosoma.
- 32. Nucleosoma.
- 33. Nucleosoma.
- 34. Nucleosoma.
- 35. Nucleosoma.
- 36. El ADN está superenrollado.
- 37. El ADN está superenrollado.
- 38. El ADN está superenrollado.
- 39. El ADN bacteriano es circular.
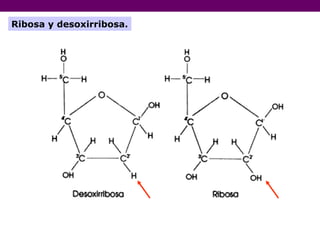
- 40. ARN. El ácido ribonucleico (ARN o RNA) es un ácido nucleico, polímero lineal de nucleótidos formando una larga cadena. El eje de la cadena lo forman grupos fosfato y azúcares ribosa de forma alternativa del que toma su nombre. Los nucleótidos del ARN contienen el azúcar ribosa y entre sus bases nitrogenadas al uracilo, a diferencia del ácido desoxirribonucleico (ADN) cuya azúcar es una desoxirribosa y contiene a la timina en vez del uracilo. La función principal del ARN es servir como intermediario de la información que lleva el ADN en forma de genes y la proteína final codificada por esos genes.
- 41. ARN. El ARN es un polirribonucleótido de ADENINA, GUANINA, CITOSINA y URACILO
- 42. Tipos de ARN.
- 43. ARNm.
- 44. ARNm.
- 45. ARNt.
- 46. ARNt.
- 47. ARNt.
- 48. Misión del ARN.
- 49. Misión del ARN.