

Recommended
More Related Content
What's hot
What's hot (20)
Viewers also liked
Similar to Ciclo de Krebs
Similar to Ciclo de Krebs (20)
Recently uploaded
Recently uploaded (20)
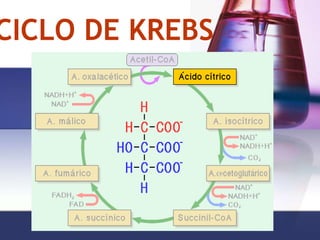
Ciclo de Krebs
- 2. HISTORIA Sir Hans Adolf Krebs o Propuso los elementos clave del consumo de O2, en cantidad desproporcionada respecto a las cantidades añadidas o Empleando malonato, lograba bloquear la oxidación del piruvato o células tratadas con malonato acumulaban citrato, succinato y α-cetoglutarato, lo cual sugería que citrato y α-cetoglutarato eran precursores del succinato
- 3. CICLO DE KREBS 1. Ácido cítrico 2. Ciclo de los ácidos tricarboxílicos 3. Ruta metabólica Sucesión de reacciones químicas, de respiración celular en las células aerobias En organismos aeróbicos, es parte de la vía catabólica que realiza la oxidación de hidratos de carbono, ácidos grasos y aminoácidos hasta producir CO2, liberando energía en forma utilizable (poder reductor y GTP).
- 4. Objetivos del Ciclo de Krebs Los objetivos del Ciclo de Krebs son: Oxidar acetil~CoA a CO2 Generar equivalentes de reducción (NADH y FADH2). Suministrar intermediarios para la síntesis de otros compuestos (Aminoácidos, Ácidos grasos, Colesterol, Gluconeogénesis, Porfirinas). Vincular derivados de aminoácidos al proceso terminal de oxidación.
- 5. Es la segunda etapa de la respiración aerobia
- 6. Dos etapas del ciclo Son 9 reacciones compartimentadas en la matriz mitocondrial catalizadas por 8 enzimas En las primeras 4 reacciones ocurren dos descarboxilaciones oxidativas, con lo cual se pierden los carbonos del acetil-CoA y se forma un compuesto activado: el succinil-CoA Las últimas 4 reacciones permiten la regeneración del oxalacetato, que hace posible que el proceso se reinicie
- 7. Origen del Acetil~CoA Por descarboxilación del piruvato. Por b oxidación de los ácidos grasos. A partir de aminoácidos cetogénicos (L, K, F, Y, I, W, T) CH3C O CoA
- 8. Fases del Ciclo de Krebs El ciclo de Krebs, como todo proceso cíclico, se inicia con la condensación de un aceptor y el abastecedor del ciclo, seguida por una serie de reacciones para eliminar el abastecedor y otras que regeneran el aceptor.
- 9. Condensación Reacciones generales del Ciclo de Krebs Oxidación Descarboxilante I Oxidación No Descarboxilante
- 11. Síntesis de citrato En la condensación participa un grupo nucleófilo en el sitio activo de la citrato sintasa, lo cual activa al carbono terminal del acetil-CoA
- 12. Isomerización del citrato La aconitasa cataliza las reacciones de deshidratación del citrato en el carbono 3 y la posterior hidratación en el carbono adyacente Esta enzima también reconoce al fluorocitrato, un sustrato suicida, cuyo producto ya no es reconocido por la siguiente enzima (isocitrato deshidrogenasa IDH)
- 14. Primera descarboxilación La isocitrato deshidrogenasa (IDH) convierte el isocitrato en α- cetoglutarato, mediante una oxidación seguida de una descarboxilación La enzima está bajo control alostérico por ADP(+) y por NADH (-)
- 15. Segunda descarboxilación El complejo α-cetoglutarato deshidrogenasa (AKGDH) oxida y descarboxila al sustrato para producir un compuesto activado: el succinil-CoA, en una reacción muy exergónica El AKGDH es similar en estructura y mecanismo de reacción al piruvato deshidrogenasa (PDH)
- 16. Síntesis de GTP/ATP Ocurre por fosforilación a nivel de sustrato Succinil-CoA + Pi + GDP <=> Succinato + GTP + CoASH (G’º = -2.9 kJ/mol) El GTP sirve como donador del fosfato al ADP para sintetizar ATP, catalizado por la nucleósido difosfocinasa (NDPK)
- 17. La oxidación del succinato dona electrones al FAD La succinato deshidrogenasa (SDH) es la única enzima del CAT unida a la membrana interna La SDH participa en otras rutas Tiene unida FAD, un aceptor de electrones que los dona directamente a la cadena respiratoria
- 18. Fumarasa La fumarato hidratasa convierte al fumarato en L-malato en una reacción estereoespecífica, que únicamente reconoce al isómero trans Fumarato + H2O <=> L-Malato
- 19. Regeneración del OAA La malato deshidrogenasa realiza la catálisis reversible de Piruvato u Oxaloacetato en Malato dependiente de NAD o NADP Mantiene los niveles de OAA intramitocondriales bajos (<1 M) L-Malato + NAD+ <=> Oxaloacetato + NADH + H+ (Gº’= +29.7 kJ/mol)
- 21. REACCIONES DEL CICLO DE KREBS
- 22. Factores que regulan la velocidad de flujo de la ruta El estado energético: la velocidad se ralentiza cuando la proporción [ATP]/[ADP] es alta La isocitrato deshidrogenasa es activada alostéricamente por el ADP Estado redox en la célula y los distintos compartimentos: la acumulación de intermediarios reducidos (FADH2 y NADH) detiene la respiración El NADH puede inhibir mediante interacciones alostéricas y por limitación del NAD+ disponible Disponibilidad de compuestos, de energía elevada y el aceptor del acetilo: varias enzimas son inhibidas por acetil- CoA y por succinil-CoA y en ausencia de oxalacetato no funciona la ruta
- 23. Los reguladores del ciclo son componentes que forman parte de un proceso más grande: la respiración
- 24. Rutas alimentadoras del ciclo de Krebs
- 25. REGULACION DEL CICLO DE KREBS DISPONIBILIDAD DE SUSTRATOS: EL CICLO DE KREBS SE ACTIVA EN: Saciedad Dietas hiperproteicas Dietas hiperlipidicas ES POCO ACTIVO EN: • Ayuno; • Ejercicio; estres; • Diabetes mellitus…
- 26. BALANCE ENERGETICO Isocitrato deshidrogenasa: NADH2 3 ATP Alfa-ceto-glutarato DHG: NADH2 3 ATP Succinato deshidrogenasa: FADH2 2 ATP Malato deshidrogenasa: NADH2 3 ATP
- 27. BALANCE ENERGETICO 11 ATP + 1 ATP (GTP) = 12 ATP En conclusión: Por cada molécula de acetil CoA que entra al ciclo de Krebs, se ganan 12 moléculas de ATP…
- 28. Destino metabólico de intermediarios del Ciclo de Krebs
- 29. DESTINO DEL CITRATO CITRATO FFQ1 ACETIL CoA COLESTEROL citoplasmática SALES BILIARES ACIDOS GRASOS VITAMINA D3 H. ESTEROIDES TRIACILGLICERIDOS MEMBRANAS
- 30. DESTINO DEL ALFA-CETO- GLUTARATO ALFA-CETO-GLUTARATO GLUTAMATO SINTESIS Neuro- GLUTAMINA PROTEICA transmision HEXOSAMINAS (GLUT,GABA) EQ.AC./BASE PURINAS UREA NH3
- 31. SUCCINIL CoA HEMO CITOCROMOS MIOGLOBINA HEMOGLOBINA CATALASA PEROXIDASA DESTINO DEL SUCCINIL CoA: PIGM.BILIARES
- 32. DESTINO DEL OXALACETATO OXALACETATO ASPARTATO PEP PROTEINAS UREA FRUCTOSA 1-6 di P PIRIMIDINAS GLUCOSA Intermedia- rios del CTC ASP, ASPn Gluconeogénesis
- 33. IMPORTANCIA DE LAS VITAMINAS EN EL CICLO RIBOFLAVINA DINUCLEÓTIDO DE FLAVINA Y ADENINA (FAD) cofactor en alfa-cetoglutarato deshidrogenasa y succinato deshidrogenasa NIACINA DINUCLEÓTIDO DE ADENINA Y NICOTINAMIDA (NAD) coenzima para isocitrato deshidrogenasa, alfa-cetoglutarato deshidrogenasa y malato deshidrogenasa. TIAMINA DIFOSFATO DE TIAMINA coenzima para la descarboxilación de alfa-cetoglutarato deshidrogenasa ACIDO PANTOTÉNICO COFACTOR adherido a residuos acilo «activo» como acetil CoA y succinil CoA