The Origin and Evolution of Sexual Reproduction
•Download as DOCX, PDF•
26 likes•12,099 views

developmental biology

Recommended
More Related Content
What's hot
What's hot (20)
Viewers also liked
Viewers also liked (20)
Similar to The Origin and Evolution of Sexual Reproduction
Similar to The Origin and Evolution of Sexual Reproduction (20)
More from Aftab Badshah
More from Aftab Badshah (20)
Recently uploaded
Recently uploaded (20)
The Origin and Evolution of Sexual Reproduction
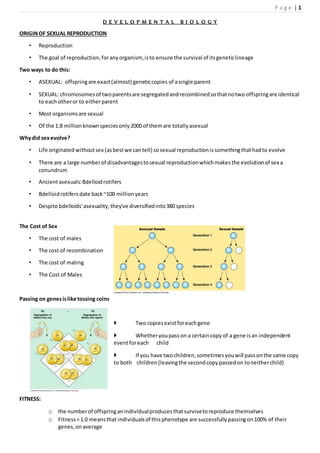
- 1. P a g e | 1 D E V E L O P M E N T A L B I O L O G Y ORIGINOF SEXUAL REPRODUCTION • Reproduction • The goal of reproduction,foranyorganism, isto ensure the survival of itsgeneticlineage Two ways to do this: • ASEXUAL: offspringare exact(almost) geneticcopies of asingle parent • SEXUAL: chromosomesof twoparentsare segregatedandrecombinedsothatnotwo offspringare identical to each otheror to eitherparent • Most organismsare sexual • Of the 1.8 millionknownspeciesonly2000 of themare totallyasexual Whydid sexevolve? • Life originatedwithoutsex (asbestwe cantell) sosexual reproductionissomethingthathadto evolve • There are a large numberof disadvantagestosexual reproductionwhichmakesthe evolutionof sex a conundrum • Ancientasexuals:Bdelloidrotifers • Bdelloidrotifersdate back~100 millionyears • Despite bdelloids'asexuality,they've diversifiedinto380 species The Cost of Sex • The cost of males • The cost of recombination • The cost of mating • The Cost of Males Passing on genesislike tossing coins Two copiesexistforeachgene Whetheryoupasson a certaincopy of a gene isan independent eventforeach child If you have twochildren,sometimesyouwill passonthe same copy to both children(leavingthe secondcopypassedon toneitherchild) FITNESS: o the numberof offspringanindividualproducesthatsurvivetoreproduce themselves o Fitness=1.0 meansthat individualsof thisphenotype are successfullypassingon100% of their genes,onaverage
- 2. P a g e | 2 How is fitnesscalculated Fitness = the numberof genespassedonto the nextgeneration Because diploidorganisms(I.e.,mostorganisms)onlypassonhalf of theirgenestoeachchild,theymust have twooffspringlivingtoreproductive age tohave Fitness=1 Fitness=1 doesnot exactly meanthat youhave passedon 100% of your genestothe nextgeneration (Remember:sometimesyousendtwocopiesof the same gene andzerocopiesof the other) Cost of recombination Asexual Sexual F F F F F F M M Fitness of females 2 1 The Cost of Mating • Cost of sexual mechanisms – Chemical attractants – Sexual organs • Cost of matingbehaviour – Courtshipiscostly – Potential exposure topredators • Disease Transmission Sexualitymust have itsadvantages • Hardlyany asexual lineagesseemold,andfossil evidencehassuggestedthatasexualityisadeadend • The prevalence of sexualityamongstspeciesiscausednotbecause asexual speciesdon'tevolve,butbecause theydon't last Red QueenHypothesis The Red QueenHypothesiswasfirstsuggestedbyLeighVanValen. Accordingto thistheory,organismshave torunfast-justtostay still! Thatis to say,theyconstantlyhave to “run to try to improve”(andthe developmentof sex wouldbe one wayof accomplishingthat). An animal constantlymustrun to chase its prey,elude predators,andresistinfectionfromdisease-causing organisms Evidence for Red QueenHypothesis In topminnows,sexual andasexuallineages coexist Sexual lineagesare the leastsusceptible toparasites Geneticvariationneededtokeepupwithevolutionof parasites Whyare babiesborn young? Somaticcellsdie,butthe germline seemstobe practicallyimmortal
- 3. P a g e | 3 In a landmarkarticle published in1989, Bernstein,Hopf,andMichodsuggestedthattheyhaddiscoveredthe answer: ‘We argue that the lackof ageingof the germ line resultsmainlyfromrepairof the geneticmaterialby meioticrecombinationduringthe formationof germcells. Muller’sRatchet hypothesis • Vastmajorityof mutationsare detrimental • Mutationacquisitionisaone-wayprocessinthe genomesof asexuals • In Salmonella typhimurium 444 lineagesstartedfromasingle colony • After1700 generations,1%of lineagesshoweddecrease infitness(growthrate) butnolineagesshowed increasedfitness Sexincreasesvariation Genesfrommaternal andpaternal parentget“shuffledup”whengametesare made Causessome gametestohave “superfit”genotypesandotherstohave “superunfit” genotypes Theory for the Originof Sexual Reproduction Viral Eukaryogenesis:Eukaryoticcellsarose fromacombinationof a DNA virusand a bacterium.The virus incorporatedgeneticmaterial fromthe bacteriumandtookoverthe role of informationstorage forthe cell.Meiosis arisesbecause of selective pressure on the virusto spreadhorizontally throughoutthe populationbycell-to-cell fusion.Twocellsinfectedwithrelated but differentvirusesfusedbecause they recognisedeachotherasuninfected. Afterthe fusionof the twocells, incompatibilitiesbetweenthe twoviruses resultina meiotic-likecell division. SPERMATOGENESIS Spermatozoadevelopfrommale primordial germcellscalledspermatogonia. Duringembryoniclife these cellsoriginateinthe endodermof the yolksacand migrate to the regionwhere testes are to develop. here theyincreaseinnumberbymitoticdivisionandfinallysettledowninthe developing seminiferoustubules. TESTES
- 4. P a g e | 4 Spermatogoniaremaindormantduringinfancyandchildhoodof the individual. Whenthe personachievespuberty,the interstitialcellsof the testesbegintosecrete testosterone,underthe influenceof thishormone the processof spermatogenesisisoccur. Withthe onsetof pubertythe spermatogoniabegin to divide mitoticallyandsoonthree varietiesof male primordial germcellscanbe identifiedinthe seminiferoustubules: (i) Type A darkspermatogonia (ii) Type A pale spermatogonia (iii) Type Bspermatogonia Type A dark spermatogonia These cellsare consideredtobe reserve cells.they occasionllydivide mitoticallytomaintaintheirown numberandto give rise to type A pale spermatogonia. Type A pale spermatogonia These cellsdividebymitosisregularlytogive rise to othertype A pale spermatogoniaaswell astype B spermatogonia. Type B spermatogonia Each type B spermatogoniumdivides mitoticallyafewtimestoproduce a numberof daughtercellswhichdonot divide furtherbutenlarge tobecome primaryspermatocytes. The spermatogoniaandthe earlyspermatocyteshave the same chromosomalconfigurationasthatof all ordinarysomaticcell. Each primaryspermatocyte replicatesitsDNA andduvidesbymeiosis-Itogive rise totwodaughtercells calledsecondaryspermatocytes. Each secondaryspermatocyte undergoesmeiosis-IItoproduce twodaughtercellscalledspermatids. The spermatidscome tolie withingroovesonthe luminal surface of sertoli cellswhere theyenterthe next stage of spermatogenesis,whichiscalledspermiogenesis.
- 5. P a g e | 5 Spermiogenesis. The processby whichspermatidsare transformedintospermatozoaiscalledspermiogenesis. Spermiogenesisinclude: condensationof nuclearchromatin Formationof acrosome Developmentof aflagellum Loss of excesscytoplasm o Duringspermiogenesisnofurtherchromosomal change buriedinthe groovesonthe surface of the sertoli cellsof the seminiferoustubulesoccurbuta greatmodificationinthe shape of the cell takesplace. o The spermatidslie buriedingroovesonthe surface of the sertoli cellsof the seminiferoustubules. o A spermatidisa small,roundcellcontainingacentrallylocatedspherical nucleus. o Importantorganellesare alsopresent: o There isa prominentgolgi apparatus o A pairof centriole o Numerousmitochondria The formation ofspermatozoon involveselaborate changesin all these cellularelements The golgi apparatusgive rise to a large vesicle,calledacrosome,whichcome tolie atthe anteriorpole of the nucleusinthe formof cap. The acrosomal material isrich incarbohydratesandcontainshydrolyticenzymes. The nucleusitself becomeprogressivelycondensed andassume ahighlyflattenedandelongatedshape. The area occupiedbythe nucleusandacrosomal cap becomeste headregionof the developing spermatozoon. As the acrosome isformng,the twocentriolesmove tothe caudal pole of the nucleus. One of the centriole give rise toflagellumwhichformthe tail of the developingspermatozoon. The cell graduallyelongates;thiselongationoccurdue to displacementof the cytoplasmcaudally. The mitochondriagraduallymove towardthe caudal regionof the cell.theareacontainingthe mitochondria isknownas middle piece of the spermatozoon. In the final stagesof the maturationof the spermatid,the residualcytoplasmispartitionedoff fromthe remainderof the spermatid. Whenthe spermis releasedfromthe seminiferousepithelium,theexcesscytoplasmbecomesdetachedfrom the spermasmembrane-boundstructure calledresidual body,whichisphagocytosedbythe sertoli cells.
- 6. P a g e | 6 The net resultof these changesis that a fullydevelopedspermatozoon i.attaina verysmall size ii.iscapable of attainingmotality Iii.attainsonlythose cellularstructures whichare essentialtomake itcapable of fertilizinganovum. OOGENESIS The processof formationanddevelopmentof ova iscalledoogenesis. Like spermatogonia,the oogoniaalsooriginate inthe endodermof the yolksacduringdevelopment. From here theymigrate toand settle downinthe developingovaries. In the ovariesthe oogoniapassthrougha phase of proliferationinwhichtheyincrease innumberbya series of mitoticdivision. Later greatnumbersof these oogoniadegenerate anddisappear. By the 12th weekof the developmentall of the persistingoogoniaenlarge insize tobecome primary oocytes,afterwhichnoproliferationoccurs.the primaryoocytesbecomesurroundedbyasingle layerof follicularcellstoformspherical orovoidstructure calledovarianfollicles. A primaryoocyte containsa relativelylarge,vesicular,eccentrallysituatednucleuswithaprominent nucleolus. The oocyte nucleusinmanyspeciesincreasessignificantlyinvolume andbecomesveryactive inRNA synthesis. Cytoplasmicorganellesare muchmore abundantinoocytes.
- 7. P a g e | 7 For instance a fullygrownXenopusoocyte contains200,000 timesasmany ribosomesasan average somatic cell. Animal eggscontainproteins,lipidsandglycogentonourishthe developingembryo.these materialswhich are collectivelycalledyolk,accumulateinthe eggcytoplasm. The amountsof yolkare minimuminmammalianeggs,whichhave tosustainembryogenesisonlytoavery immature stage. In contrastthe large eggsof birdsand reptileshave large amountof yolktosupportembryonicdevelopment to advancedstages. In the course of embryogenesis,yolkisbrokendownintosmall molecules,suchasaminoacids,whichare then usedbyembyoniccellsintheirsyntheticprocesses. Like RNA,yolkaccumulatesinthe oocyte duringmeioticarrestandcontributestothe oocyte’senormous growth. Major egg proteinsare calledvittelins,whichare formedfromprecursorcalledvitellogenins. In vertebratesvitellogeninsynthesisoccurinliver. In insectsthe majorproductionsite isthe fatbody. Initiallythe plasmalemmaof the oocyte issmoothandinclose appositionwiththe surroundingfollicular cells.however,asthe oocyte enlargesandthe zonapellucidaformsaroundit,auniformcoverof microvilli developsoverthe oolemma. The golgi apparatusproducesa special type of granulescalledcortical granules.theyare boundedbyaunit membrane andcontainenzymeswhichplayaveryimportantrole inblockingpolyspermy. At the endof the growthperiod,all the primaryoocytesreplicate theirDNA andthenenterthe prophase of mieosis-I,butthe completionof the prophase of thisdivisiondoesnotoccurdurinngthe restof the intra- uterine life.duringthisperiodandfora longtime afterbirth,thesecellsremainarrestedin the prophase of meiosis-I.thisprolongedperiodof suspendedprophase persistsforseveral years. Eventually,withthe onsetof pubertythe processof oogenesisisreactivated. Duringthe follicularphase of eachovariancycle manyovarianfolliclesgrow underthe influence of FSHbut onlyone of themovulates,othersdegenerates. Shortlybefore ovulation,the primaryoocytecompletesmeiosis-I anddivide intotwodaughtercells. Unlike spermatogenesis,thedivisionof cytoplasmismarkedlyunequal. one of the daughtercellsrecievesmostof the cytoplasmandisknownas secondaryoocyte;the othercell calledfirstpolarbodyrecievesverylittlecytoplasm.itisasmall non-functionalcell whichmaydivide again but eventuallyundergoesdegeneration. As soonas meiosis-Iiscompleted,the secondaryoocyte entersthe meiosis-IIbutthe divisionprogressesonly uptothe metaphase whenthe processisonce againarrested.thissocalledovumisreleasedatovulation. The secondmeioticdivisionwill onlybe completedif the ovumisfertilizedbythe sperm. Eggs of differentanimal speciesare ovulatedandfertilizedatdifferentstagesof mieosis. As soonas the cell membrane of the ovumispenetratedbyasperm,the oocyte completesthe second meioticdivisionandtwodaughtercellsare produced.once again,mostof the cytoplasmgoestoone cell whichiscalledootid.the othercell calledsecondpolarbody,issmallandfuctionanlessthatsoon degenerates. Eggs are protectedbyelaborate envelopes. The plasmamembrane of eggcell iscoveredbya glycoproteinlayer,calledzonapellucidainmammalsand generallyreferredtoasvitelline envelope whichplays animportantrole infertilization.
- 8. P a g e | 8 In contrast,manytypesof eggsdepositedonland,suchasthose of reptilesandbirds,have hardshells.for instance the yolkychickeneggissurroundedinitiallybyafragile vitellineenvelope.above thisenvelope several layersof “egg white”consistedof ovalbuminisdepositedabovewhichshell membranesare added. Fertilization Fertilizationisthe processwherebytwosex cells(gametes) fusetogethertocreate a new individual . Fertilizationaccomplishestwoseparate ends:sex (thecombiningof genesderivedfromthe twoparents) and reproduction(the creationof neworganisms). Thus, the firstfunctionof fertilizationistotransmitgenesfromparenttooffspring,andthe secondisto initiate inthe eggcytoplasmthose reactionsthatpermitdevelopmenttoproceed. Althoughthe detailsof fertilizationvaryfromspeciestospecies,conceptiongenerally consistsof fourmajor events 1. Recognitionbetweenspermandegg. 2. Regulationof spermentryintothe egg. 3. Fusionof the geneticmaterial of spermandegg. 4. Activationof eggmetabolismtostartdevelopment.
- 9. P a g e | 9 Recognitionof spermand egg The interactionof spermandegg generallyproceedsaccordingtofive basicsteps 1. The chemoattractionof the spermto the eggby soluble moleculessecretedbythe egg 2. The exocytosisof the acrosomal vesicle toreleaseitsenzymes 3. The bindingof the spermtothe extracellularenvelope(vitelline layerorzonapellucida) of the egg 4. The passingof the spermthroughthisextracellularenvelope 5. Fusionof egg and spermcell plasmamembranes Enclosingthe cytoplasmisthe egg plasma membrane.Outside the plasmamembrane isthe vitelline envelope.Thisenvelopecontainsglycoproteinsandisofteninvolvedinsperm-eggrecognition.Itis supplementedbyextensionsof membrane glycoproteinsfromthe plasmamembrane andbyproteinaceous vitelline poststhatadhere the vitelline envelopetothe membrane.The vitellineenvelope isessential forthe species-specificbindingof sperm. In mammals,the vitellineenvelope isaseparate andthickextracellularmatrix calledthe zonapellucida. The mammalianeggisalso surroundedbyalayerof cellscalledthe cumulus,which ismade up of the ovarianfollicularcellsthatwere nurturingthe eggatthe time of itsrelease fromthe ovary.Mammalian spermhave to getpast these cellstofertilize the egg.The innermostlayerof cumuluscells,immediately adjacentto the zona pellucida,iscalledthe coronaradiata. External fertilizationinseaurchins Many marine organismsrelease theirgametesintothe environment. These organismsare facedwithtwoproblems:How can spermandeggsmeetinsuch a dilute concentration, and howcan spermbe preventedfromtryingtofertilizeeggsof anotherspecies? Two majormechanismshave evolvedtosolve theseproblems: speciesspecificattractionof spermandspecies-specificspermactivation. Sperm attraction: One chemotacticmolecule,called resact,hasbeenisolatedfrom the eggjellyof the seaurchin Arbacia punctulata . Resact diffusesreadily in seawaterandhasa profoundeffectatverylow concentrationswhenaddedtoa suspensionof Arbacia sperms. Resact isspecificforA.punctulata and doesnotattractspermof other species. A.punctulata spermhave receptorsin theirplasmamembranesthatbindresact and can swimupa concentrationgradientof this compounduntil theyreachthe egg. The acrosomal reaction in sea urchins Acrosomal reactioninseaurchinsis initiatedbycontactof the spermwiththe eggjelly. In sea urchins,the acrosomal reactionisthoughttobe initiatedbyafucose-containingpolysaccharide inthe eggjellythatbindstothe spermandallowscalciumto enterintothe spermhead. In most marine invertebrates,the acrosomal reactionhastwocomponents: the fusionof the acrosomal vesicle withthe spermplasmamembrane and the extensionof the acrosomal process
- 10. P a g e | 10 Species-specificrecognitioninseaurchins Once the sea urchinspermhas penetratedthe eggjelly,the acrosomal processof the spermcontactsthe surface of the egg. A majorspecies-specificrecognitionstepoccursat thispoint.The acrosomal protein mediatingthisrecognitioniscalled bindin. it iscapable of bindingtodejelliedeggsof the same species. Thus,species-specificrecognitionof seaurchingametesoccursat the levelsof spermattraction,sperm activation,andspermadhesiontothe eggsurface. Once the sea urchinsperm has penetratedthe eggjelly,the acrosomal processof the spermcontactsthe surface of the egg. Gamete Fusionand the PreventionofPolyspermy 1) Fusionof the eggand spermplasma membranes o Recognitionof spermbythe vitellineenvelope is followedbythe lysisof thatportionof the envelope of the spermheadby the acrosomal enzyme.Thislysisisfollowedbythe fusionof the spermplasmamembrane withthe plasmamembrane of the egg. 2) The preventionofpolyspermy o The fast blockto poly-spermyisachievedby changingthe electricpotential of the eggplasmamembrane. o Thismembrane providesaselective barrierbetweenthe eggcytoplasmandthe outside environment,and the ionicconcentrationof the eggdiffersgreatlyfromthatof itssurroundings. o Within1 3 secondsafterthe bindingof the firstsperm, the membrane potentialshiftstoapositive level, about+20 mV . This change iscausedby a small influx of sodiumionsintothe egg. 3) Slow block to polyspermy o The slowblock to polyspermy. o Directlybeneaththe seaurchineggplasmamembrane are about15,000 cortical granules. o Upon spermentry,these cortical granulesfuse withthe eggplasmamembrane andreleasetheircontents intothe space betweenthe plasmamembrane andthe vitelline envelope proteins.
- 11. P a g e | 11 Several proteinsare releasedbythiscortical granule exocytosis.The firstare proteases.Theseenzymes dissolve the proteinpoststhatconnectthe vitellineenvelope proteinstothe cell membrane,andtheyclip off the bindinreceptorandanyspermattachedto it . Mucopolysaccharidesreleasedbythe cortical granulesproduce anosmoticgradientthatcauses waterto rush intothe space betweenthe plasmamembraneandthe vitelline envelope,causingthe envelopetoexpandandbecome the fertilizationenvelope.A thirdproteinreleasedbythe cortical granules, a peroxidase enzyme,hardensthe fertilizationenvelope. Finally,afourthcortical granule protein,hyalin,formsacoatingaroundthe egg . The egg extendselongatedmicrovilli whosetipsattachto this hyaline layer.This layer providessupport for the blastomeresduringcleavage. Fusionof genetcmaterial In seaurchins,the spermnucleusentersthe eggperpendiculartothe eggsurface.Afterfusionof the sperm and eggplasmamembranes,the spermnucleusanditscentriole separate fromthe mitochondriaandthe flagellum.The mitochondriaandthe flagellumdisintegrate insidethe egg,soveryfew,if any,sperm-derived mitochondriaare foundindevelopingoradultorganisms. Thus,althougheachgamete contributesahaploidgenome tothe zygote,the mitochondrial genome is transmittedprimarilybythe maternal parent. Afterthe seaurchinspermentersthe eggcytoplasm, the male pronucleusrotates180° sothat the sperm centriole is betweenthe spermpronucleusandthe eggpronucleus.The spermcentriole thenactsasa microtubule organizingcenter,extendingitsownmicrotubulesandintegratingthemwitheggmicrotubules to forman aster. These microtubulesextendthroughoutthe egg andcontact the female pronucleus,andthe twopronuclei migrate towardeach other. Theirfusionformsthe diploid zygote nucleus. The Activation ofEgg Metabolism Althoughfertilizationisoftendepictedasmerelythe meanstomerge twohaploidnuclei,it hasanequally importantrole ininitiatingthe processesthatbegindevelopment.These eventshappenincytoplasmand occur withoutthe involvementof the parental nuclei. The release of calciumionsthatoccurs whenthe spermentersthe eggiscritical for activatingthe egg’s metabolismandinitiatingdevelopment. Calciumionsrelease the inhibitorsfrommaternallystoredmessages,allowingthese mRNAstobe translated. Theyalsorelease the inhibitionof nucleardivision,therebyallowingcleavagetooccur. IP3 Pathway The membrane phospoholipidphosphotidylinositol4,5-bisphosphate issplitbythe enzyme phospholipaseC to yieldtwoactive compounds i.e IP3(inositol 1,4,5-triphosphate)andDAG(diacylglycerol). IP3 isable to release calciumionsintocytoplasmbyopeningthe calciumchannelsof endoplasmicreticulum. DAG activatesproteinkinase Cwhichinturnactivate Na+/H+exchange pump. These processesresultinthe liberationof Ca2+ and the alkalinizationof eggandtherefore areinvolvedinthe developmentof egg.
- 12. P a g e | 12 CLASSIFICATIONOF EGGS The Structure of a typical Ovum • Ovumis the female gamete.ltstoresfoodrequiredforthe entireprocessof developmentinthe formof yolk.lthas three importantf unctions: 1. lt suppliesahaploidsetof chromosomestothe future embryo. 2. lt contributesalmostall cytoplasmtothe zygote. 3. lt suppliesfoodtothe developingembryo Shape and Size • Typically,the eggsare spherical orovoidinshape.Butina few animalslike insects,the eggsare elongated and cylindrical .Eggsare generallylargerthanthe spermsandaverage somaticcells. The egg iscoveredexternallybyaplasmamembrane orplasmalemma.Withinthe plasmamembraneisthe granularcytoplasm. Organization of Egg Cytoplasm • The cytoplasmof egg is knownasooplasm.ltisgranular andcontainsin additiontothe usual cellular organellescertainotherinclusionsIikeyolkandcortical granules.The peripherallayerof ooplasmismore viscousandgelatinous.ltis knownasthe eggcortex whichis providedwithmanymicrovilli andcortical granules.The microvilli are formedbythe outpushingsof the plasmalemmaandtheyhelpintransportation of substancesfromthe outside intothe ooplasmduringthe developmentof egg. • Theyare membrane boundandare formed fromgolgi complex.Theycontainhomogeneousandgranular polysaccharides.Cortical granulesare presentinthe eggsof seaurchins,frogs,fishes,bivalvemolluscs, several annelidsandcertainmammals. Yolk: • Nutritive substancesare storedinthe cytoplasmof egginthe form of yolk. Thisstoredfoodis utilizedbythe embryoforits earlydevelopment.The processof formationof yolkisknownasvitellogenesis.The yolkisa complex material consistingof proteins,fats,carbohydratesetc.The yolkmaybe called"proteinyolk"when it hasmore proteinsthanlipids,or" fattyyolk"whenithas more fat contentsthanthe proteins.Most animal eggscontainbothkindsof yolk. ClassificationofEgg • 1. On the Basis of the Amountof yolk • 2. On the Basis of the distribution of yolk
- 13. P a g e | 13 • 3. On the basisof potentialitiesof eggforfurtherdevelopment Classificationofegg on the basisof amount of yolk • Eggs are groupedintofourtypesonthe basisof the amountof yolk presentinthem. a. Alecithal egg b.Microlecithal egg c.Mesolecithal egg d.Macrolecithal egg 1) Alecithal egg: Whenthe egg contains negligibleamountof yolk,itis calledalecithal egg. Eg. The eggsof Eutherianmammals 2)Microlecithal egg: Whenthe egg containsmall amountof yolkit issaidto be microlecithal. Romerand Balinskynamedthese eggsasoligolecithal eggs. Eg.Amphioxus,Tunicates 3)Mesolecithal egg: Whenthe amountof yolkpresentismoderate andisnothigh,these eggsare alsonamedas mesolecithal eggs e.gamphibian,Dipnoi ,xenopus andPetromyzon 4)Macrolecithal egg: Whenthe egg containslarge amountof yolkit issaidto be macrolecithal ormegalecithal egg. It isalso calledPolylecithal egg. Eg. bonyfishes'amphibians,reptilesandbirds. Etc Alecithal egg microlecithal egg mesolecithal egg macrolecithal egg On the Basis of the distribution of yolk a. Isolecithal egg b. Telolecithal egg c. Centrolecithal egg a. Isolecithal orHomolecithal Egg: lnisolecithal eggs,the verylittleamountof yolkpresentisuniformlydistributedthroughoutthe ooplasm (e.gechinoderms,Amphioxus,mammals).Thisconditionisusuallyobservedineggswithverylittleamount of yolk.
- 14. P a g e | 14 b. Telolecithal Egg: In eggscontainingmoderate orlarge quantityof yolk,the distributionof yolkisnotuniform.ltis concentratedmore towardsthe vegetal pole.Suchatype of egg,in whichthe yolkisconcentratedtowards one pole,is calledtelolecithal egg. Telolecithal eggsmayfurtherclassifiedintothree types 1. SlightlyTelolecithal This type of egg containsonlya small quantityof yolkwhich isdistributedunevenly. The vegetal pole hasthe highestconcentrationandthe animal pole the lower(e.g.eggsof fishes). 2. ModeratelyTelolecithal egg Thistype of eggcontainsa moderate quanilty of yolkwhichis Distributed unevenly.Due tohighconcenteration of yolk inthe vegetal hemisphere,the nucleusis shiftedmore towardsthe animal hemisphere (eg.amphibianegg). 3. ExtremelyTelolecithalEgg lnthistype of egg,due to the heavydepositionof yolk,the entire vegetal hemisphereandamajor portionof the animal hemisphere are occupiedbyyolk.Due tothisextremely unevendistributionof yolk,the ooplasmandnucleusare displacedtowardsthe animal pole(e..g reptilianandavianeggs). c. Centrolecithal egg: • Egg of manyarthropodsand some coelenteratesare describedascentrolecithal.Theyare relativelylarge and elongate andhave averygreat amountof yolk.The nucleusliesatthe geometriccentre of the yolk mass,surroundedbya small amountof cytoplasm.A thincytoplasmiclayercoversthe surface of the yolk. Fine strandsof cytoplasmextendfromthe peripheral layertothe zone occupiedbythe nucleus. On the basis ofpotentialitiesforfurther development 1)Mosaic Egg: ln certaineggs,everyportionispredeterminedwithrespecttoitspotentialitiesforfurtherdevelopment lf a small portionof suchan eggis removed,adefectiveembryoisformed,Thisisbecause removalof aportion resultsina permanentlossfromthe egg.The remainingportionof the eggcannotmake compensatorydevelopment for the lostpart. Suchan egg, iscalledmosaicor determinateegg(e.gannelids,Molluscs). 2)Regulative Egg lnvertebratesandmostof the invertebrates,the developmentalpotentialitiesare notpredeterminedinthe eggs. Removal of a small portionof the egg,or evenone or twoearlyblastomereswill notaffectthe normal development. Thistype of eggin whichthe future developmentalpotentialitiesare notpredeterminedisknownasregulativeor indeterminate egg.
- 15. P a g e | 15 Cleavage Cleavage isarapid seriesof mitoticdivisionsthatoccur justafterfertilization. There are twocritical reasonswhycleavage issoimportant: Generationof a large numberof cellsthatcan undergodifferentiationandgastrulationtoformorgans. Increase inthe nucleus/cytoplasmicratio.Eggsneeda lotof cytoplasmtosupportembryogenesis.Itis difficultorimpossibleforone nucleustosupportahuge cytoplasm, andoocytesare one of the largestcells that exist.One small nucleusjustcannottranscribe enoughRNA tomeetthe needsof the huge cytoplasm. A largernucleustocytoplasmicratioisoptimal forcell function.Celldivision occursrapidlyafterfertilization to correct thisproblem. Cleavage differsfromnormal mitosesin 2 respects 1. Blastomeresdonotgrowin size betweensuccessivecell divisionsastheydoinmostcells.Thisleadstoa rapidincrease inthe nucleus/cytoplasmicratio.Cellsundergoingcleavagehave mainlySandM phasesof the cell cycle (little ornoG1 or G2). 2. Cleavage occursveryrapidly,andmitosisandcytokinesisineachroundof cell divisionare completewithin an hour.Typical somaticcellsdivide muchmore slowly(several hourstodays) andeventhe fastestcancer cellsdivide muchslowerthanoccursin a zygote duringcleavage. Cleavage differsindifferenttypesof eggs.The presence of large amountsof yolkaltersthe cleavagepattern,leading to incomplete cleavage thatcharacterizesbirdsandreptiles. Eggs are classifiedby how much yolk ispresent 1. Isolecithal eggs(iso=equal) have asmall amountof yolkthatis equallydistributedinthe cytoplasm(most mammalshave isolecithal eggs). 2. Mesolecithal eggs(meso=middle) have amoderate amountof yolk,andthe yolkispresentmainlyinthe vegetal hemisphere (amphibianshave mesolecithal eggs). 3. Telolecithal eggs(telo=end) have a large amountof yolkthatfillsthe cytoplasm, exceptforasmall area nearthe animal pole (fish,reptiles,andbirds). 4. Centrolecithal eggshave alotof yolkthatis concentratedwithinthe centerof the cell (insectsand arthropods). The pattern of cleavage of the zygote dependsuponthe pattern of yolk distribution 1. Holoblasticcleavage:occursinisolecithal eggs(mammals,seaurchins).The entire egg iscleavedduringeachdivision. 2. Meroblasticcleavage occurswheneggs have a lot of yolk.The egg doesnotdivide completelyateachdivision.Twotypes: Discoidal cleavage islimitedtoa small discof cytoplasmat the animal pole.All of the yolkfilledcytoplasmfailsto cleave (characteristicof telolecithal eggssuch as birds). Superficial cleavage islimitedtoa thin surface area of cytoplasmthatcovers the entire egg.The inside of the egg that isfilledwithyolkfails tocleave (centrolecithal eggssuchasinsects).
- 16. P a g e | 16 Sea UrchinsHave Isolecithal Eggs And UndergoHoloblastic Cleavage Cleavage plane:thisisthe plane inwhichcleavageoccurs.Itis orientedatrightanglestothe metaphase plate.Inseaurchins,the firstcleavage ismeridional. Meridional cleavage runsfromone pole toanother(toptobottom),like the meridianonaglobe. The secondcleavage isalsomeridional. Equatorial cleavage encirclesthe zygote like the equatoronthe globe.The thirdcleavage inthe seaurchinis equatorial.Thiscreatesananimal andvegetal half. The fourthcleavage isunique.Equal cytokinesisoccursinthe four blastomeres of the animal pole,givingrise to 8 mesomeres(allthe same size). Unequal cytokinesisoccursinthe vegetal pole.Thiscauses4large macromeresand4 small micromeres The 5th cleavage ismeridional.All mesomeresdivideequallyasdothe macromeres. As cleavage progresses,all blastomeresadhere atthe outersurface,butattachmentislostat the inner surface.The blastocoel isacavityformeddue to the unequal adherence of blastomeres. Amphibianeggs have moderate amount of yolk,however,theyare still able toundergoholoblasticcleavage. The 1st cleavage ismeridional,asisthe 2nd.The 3rd cleavage isequatorial.The cleavageisdisplacedtowardthe animal pole due tothe yolk.Thisresultsin4 small animal blastomeresand4 large vegetal blastomeres. Morula (morum= mulberry) atthe 16 to 32 cell stage the embryoiscalledamorulabecause itlookslike amulberry.
- 17. P a g e | 17 The blastocoel is displacedto the animal pole in amphibians Blastula= fromthe 128 cell stage onwardthe amphibianembryois ablastula.The outersurface of the amphibianblastulahascellsconnectedby specializedcell junctions. Tightjunctionscreate a seal thatisolatesthe outside of an embryofromthe innerlayer.Tightjunctions polarize the apical andbasal surfaces.The basal portionsof cellsstartsecretingintothe blastocoel. Desmosomesattachthe blastomerestogetheronthe outside. Gap junctionsconnectall surface blastomeres. Mammalian eggshave rotational cleavage that is holoblastic The mammalianeggisa little slow.Itbeginstocleave in the oviductand continues until itimplantsinthe wall of the uterus(1 cleavage /24 hr). Asynchronouscleavage: mammalianembryosare unusual inthat theyhave asynchronouscleavage.Not all blastomeresdivideatthe same time. The firstcleavage ismeridional,andthe secondcleavage isrotational.The 2 blastomeresdivideindifferent planes(one isequatorial andone ismeridional. Mammalian embryosundergo compaction at the 8 cell stage At first,the blastomeresof mammalianembryoshave aloose arrangement,andtouchonlyatthe basal surfaces. Aftercompaction,blastomeresadheretightly,maximizingthe areaof contact. Duringcompaction,eachblastomere undergoespolarization.Tightjunctionsdeveloponthe outersurface,
- 18. P a g e | 18 allowingproteinstospecialize.Cellstake upfluidsfromthe uterine environmentand Secrete intothe blastocoel. Gap junctionsformonthe outercellstoaidin intercellularcommunication. A blastocoel developsas cleavage proceedsto the 32-64 cell stage Aftercompactionat the 8-16 cell stage,there are 2 typesof blastomeres.Outsideblastomeresare tightly joinedandnumberabout9-14. Theysurround2-7 inside blastomeres thatare looselyjoined. Cavitation:the outside blastomeresstarttotake up fluidfromthe uterusandpumpit intothe center, creatingthe blastocoel.The blastocystisthe characteristicof earlyembryonicdevelopmentinmammals. Innercell mass:this givesrise tothe embryo,anddevelopsfromthe insideblastomeres Trophoblast:a structure consistingof outside blastomeres,thiscontributestoformingthe placenta. Embryonic stem cellscan be culturedfrom the innercell mass Cellsinthe innercell mass are undifferentiated,they multiplyindefinitely,andare knownas embryonicstem cells.Stemcellsare totipotent=theyhave the potential toformanytissue. These cellsare of great scientificandmedical importance. Theycan be removedfrom the embryo,genescanbe introducedintothe cells,and thentheycan be placedback in the blastocyst.Thisishowone constructstransgenicor “knock out” mice. The embryonicstemcellsare alsousedto grow certaintypes of tissue inculture. Theoretically,itshouldbe possible togrowstructures such as ears,muscles,nerves, and skinfortransplantationto sickindividuals. Interestingly,if youinject adult,differentiatedcellsback intothe environmentof the morulaor blastula,they become undifferentiated,and theycan redifferentiate to formmany parts of the body. Developmentofmonozygoticor identical twins Monozygotictwinsdevelopfromone zygote bysplittingatvariousstagesof development(fromthe 2cell to the blastocyststage). The stage of splittingeffectsthe overallstructure of the embryoandextra embryonicmembranes.Conjoinedtwins • are identical twinswhodevelopfromasingle fertilizedovum. • are alwaysthe same sex and race.
- 19. P a g e | 19 • are more oftenfemale thanmale,ataratio of 3:1. • occur once in 40,000 births but onlyonce in200,000 live births. • maybe causedbyany numberof factors,beinginfluencedbygeneticandenvironmentalconditions. Gastrulation in mammals Mammalian eggshave rotational cleavage that is holoblastic The mammalianeggisa little slow.Itbeginstocleave in the oviductand continuesuntil itimplantsinthe wall of the uterus(1 cleavage /24 hr). Asynchronouscleavage:mammalianembryosare unusual inthat theyhave asynchronouscleavage.Notall blastomeresdivideatthe same time. The firstcleavage ismeridional,andthe secondcleavage isrotational.The 2 blastomeresdivideindifferentplanes (one isequatorial andone ismeridional. Mammalian embryosundergo compaction at the 8 cell stage At first,the blastomeresof mammalianembryoshave aloose arrangement,and touch onlyat the basal surfaces.
- 20. P a g e | 20 Aftercompaction,blastomeresadheretightly,maximizingthe areaof contact.Duringcompaction,eachblastomere undergoespolarization. Tightjunctionsdeveloponthe outersurface,allowingproteinstospecialize. Cellstake upfluidsfromthe uterine environmentand Secrete intothe blastocoel. Gap junctionsformonthe outercellstoaidin intercellularcommunication. A blastocoel developsas cleavage proceedsto the 32-64 cell stage Aftercompactionat the 8-16 cell stage,there are 2 typesof blastomeres.Outside blastomeresare tightly joinedandnumberabout9-14. Theysurround2-7 inside blastomeresthatare looselyjoined. Cavitation:the outside blastomeresstarttotake up fluidfromthe uterusand pumpit intothe center,creatingthe blastocoel.The blastocystisthe characteristicof earlyembryonicdevelopmentinmammals. Innercell mass:thisgivesrise tothe embryo,anddevelopsfromthe inside blastomeres Trophoblast:a structure consistingof outside blastomeres,thiscontributesto formingthe placenta. Beginningof implantation The blastocystliesfree inthe uterine secretionsforabout2 days. On the 6th dayof developmentthe zonapellucidadisappearsandthe conceptusbeginstoimplantinto endometrium. The trophoblastbegintodifferentiate intotwolayers i.the outerlayercalledsyncytiotrophoblast, consistsof multinucleatedprotoplasmicmassinwhichnocell boundariescanbe distinguished Ii .the innerlayer, termedcytotrophoblast, ismade upof mono-nucleatedcellswithdistinctboundaries The syncytiotrophoblastpossessactive invasivepropertiesbecause itsecrete hydrolyticenzymes Completionofimplantation On day7 the blastocystbecome tightlyadherenttothe epitheliumof the endometriumdue toinvasive activityof the syncytiotrophoblast. The cells of endometriumrespondbydifferentiatingintolarge polyhedral cells,whichare ladenwithlipidsand glycogen. The endometriumisnowreferredtoasdecidua. As implantationprogresses,the expandingsyncytiotrophoblastgraduallyspreadsoverthe outer surface of the blastocystandby day9 the blastocystisalmostcompletelycoveredbyathicklayerof syncytiotrophoblast. Formation of bilaminargerm disc Withthe beginningof the secondweek,the cellsof the embryoblastbecome differentiatedintotwo layers, Epiblastand hypoblast Formationof amnioticcavity Developmentof primaryyolksac
- 21. P a g e | 21 Gastrulation Gastrulationinmammalsbeginswiththe appearance of primitivestreakonthe dorsal surface of the embryonicdisc On day15 of developmentthe primitive streakcanbe seeninthe longitudinal midline of the caudal partof the embryonicdiscasa narrowgroove withslightlyelevatedmargins. These elevationsare producedbyproliferationandaccumulationof epiblasticcells. The groove withinthe primitivestreakiscalledprimitive groove. By the 16th day a small circularswellingappearsatthe cranial endof the primitive streak.Thisswellingcalled primitive node,showsasmall depressioncalledprimitive pit. At itscranial end,the primitive groove become continuouswiththe primitive pit. Afterformationof the primitivestreakandappearance of the primitivegroove,the epiblasticcellssurroundingthe primitive streakbegintoproliferate,flattenandlose theirconnectionswitheachother. These epiblasticcellsmigrate towardsthe primitive streak As the primitive grooveisarrived,thesecellsslipthroughthe groove intospace betweenthe epiblastand hypoblast.thisinwardmovementof the epiblasticcellsiscalledinvagination. Those epiblasticcellswhichenterthe cranial partof the primitive streakinvade the hypoblastanddisplace itscells, so that eventuallythe hypoblastbecomescompletelyreplacedbyanew layerof cellscalleddefinitive endoderm. The definitive endodermformsthe roof of the yolksac. Otherepiblasticcellswhichinvaginatethroughthe primitive streakcome tolie betweenthe definitive endoderm and the epiblast.These cellsspreadlaterallyandforwardstoforma new layerknownasembryonicmesodermalso calledintraembryonicmesoderm. Afterformationof the embryonicmesoderm, thoseepiblastic cellswhichremainonthe dorsal surface of the embryonicdisc nowconstitute embryonicectoderm. Under the embryonicmesodermliesthe definitive endoderm whichisnowreferredtoas embryonicendodermorsimply endoderm. Thus,the two-layered(bilaminar) embryonicdisc(consisting of epiblastandhypoblast) hasnowbecome three-layered(trilaminar). The three layersof the trilaminarembryonicdisc(ectoderm, mesodermandendoderm) are calledprimarygerm layers.All tissuesandorgansof the humanbodywill derive fromthesethree layers.
- 22. P a g e | 22 Formation of notochord Developmentof notochordconsistsof twoevents 1. Formationof notochordal process 2. Formationof notochordfrom the notochordal process Formation of notochordal process The epiblasticcells,whichenterthe primitive pitgive rise toamesodermal tube betweenthe ectodermand endoderm;thistube iscallednotochordal process. The tubularnotochordal processgrowsinlengthas the cellsproliferatinginthe regionof the primitivenode are addedat the proximal endof the tubularprocess. Formation of notochord By the 18th day of developmentthe floorof the notochordal processfuseswiththe underlyingendoderm.The fused layersoondegenerate anddisappear,resultinginthe lossof floorof the notochordal process. The openingatthe primitive pitnowtemporarilyconnectsthe amnioticcavityandyolksac,this communicationis calledneuroentericcanal Afterthe disappearance of the floor,the roof of the notochordal processformsaflattenedmid-ventral barof mesoderm,callednotochordal plate,whichliesinthe roof of the yolksac intercalatedbetweenthe endodermal cells.Lateron the notochordal plate detachesfromthe endodermandgraduallyrollsuptoforma soldchord of cells, whichiscallednotochord.
- 23. P a g e | 23 Whenthishappensthe endoderminthe roof of the yolksac becomes continuous once againandnogap remainsin the endodermal liningof the yolk sacroof. Neurulation Duringthe 3rd weekof development,the notochordinducesthe overlyingectodermtodifferentiateintoathickplate of pseudo-stratifiedcolumnarneuro-epithelial cells. Thisplate iscalledneural plate andthe ectodermformingthis plate isknownas neuroectoderm. The restof the ectodermwhichliesaroundthe neural plate isnow referredtoas surface ectoderm. By the endof the 3rd weekof development,the neural plateinvaginatesalongitscentral axistoforma groove called neural groove.The foldsformingthe sidesof the groove are termedneural folds.The edge of eachfoldisknownas neural crest. Duringthe 4th weekthe neural groove deepens,the neuralfoldsbecome concave androtate aroundthe neural groove so thattheirlateral lipsmeetdorsallytoformatubularstructure,whichiscalledneural tube. As the lipsof the neural tube fuse,the junctionbetweenthe neuroectoermandthe adjacentsurface ectodermis pulleddorsallyandthe opposingmarginsof the surface ectodermalsomeetandfuse.Due tothisfusion,the neural tube separatesfromthe surface ectoderm. Neurulation Neurulation is a part of organogenesis in vertebrate embryos. Stepsof neurulationincludethe formationof the dorsalnervecord,andthe eventualformationof the centralnervous system. The process begins when the notochord induces the formationof the central nervous system (CNS) by signaling the ectoderm germ layer above it to form the thick and flat neural plate.
- 24. P a g e | 24 The neural plate foldsinuponitself toformthe neural tube,whichwill laterdifferentiate intothe spinalcord andthe brain, eventually forming the central nervous system. Primary neurulation Induction Primary neurulation occurs in response to soluble growth factors secreted by the notochord.Ectodermal cellsare induced to form neuroectoderm from a variety of signals.Ectodermsendsandreceivessignals of BMP4 (bone morphogenic protein) and cellswhichreceiveBMP4signal developinto epidermis. The inhibitory signals chordin, noggin and follistatin are needed to form neural plate. These inhibitory signals are created and emitted by the notochord. Cells which do not receive BMP4 signaling due to the effects of the inhibitory signals will develop intothe anteriorneuroectodermcellsof the neural plate. Shape Change The cells of the neural plate are signaledto become high-columnarandcanbe identified through microscopy as different from the surrounding epiblastic ectoderm. The cells move laterally and away from the central axis and change into a truncated pyramid shape. Folding The process of the flat neural plate folding into the cylindrical neural tube is termed primary neurulation As a result of the cellular shape changes, the neural plate forms the medial hinge point (MHP). The expanding epidermisputspressure onthe MHP and causesthe neural plate to foldresultingin neural foldsand the creationof the neural groove. The neural folds meet and fuse at the midline. The folding of the neural tube to form an actual tube does not occur all at once. The lateral edgesof the neuralplate touchinthe midline andjointogether.Thiscontinuesbothcraniallyandcaudally. The openings that are formed at the cranial and caudal regions are termed the cranial and caudal neuropores. In human embryos, the cranial neuropore closes approximately on day 25 and the caudal neuropore on day 27.
- 25. P a g e | 25 Secondary Neurulation In secondary neurulation, the neural ectoderm and some cells from the endoderm form the medullary cord. The medullary cord condenses, separates and then forms cavities. These cavities then merge to form a single tube. Secondary Neurulation occurs in the posterior section of mostanimalsbutitisbetterexpressedinbirds.Tubesfrom both primary and secondary neurulation eventually connect. Early brain development The anterior segment of the neural tube forms the three main parts of the brain: the forebrain, midbrain, and the hindbrain. Formation of these structures begins with a swelling of the neural tube. Ion pumps are used to increase the fluid pressure within the tube and create a bulge.A blockage betweenthe brainandthe spinalcordpreventsthe fluidaccumulationfromleakingout. Thesebrainregions furtherdivideintosub-regions.Theneuraltube becomesthe germinalneuro-epitheliumandservesasasource of new neurons during brain development. Non-neural ectoderm tissue Mesoderm surrounding the notochord at the sides will develop into the somites (future muscles, bones, and contributes to the formation of limbs of the vertebrate). Neural crest cells Masses of tissue called the neural crest that are located at the very edges of the lateral plates of the folding neural tube separate from the neural tube and migrate to become a variety of cell populations, including the cells of the peripheral nervous system.