More Related Content
Similar to 3 gen 2 76 (8)
3 gen 2 76
- 1. Mutation&DNA repair
การกลายพันธุ์(Mutation) หมายถึง ภาวะทีมีการเปลียนแปลงของสารพันธุกรรม ทําให้สิงมีชีวิตมีลักษณะแตกต่างจาก
เดิม ข้อดีของmutation คือ การวิวัฒนาการ/การกลายพันธ์ทําให้เกิดสิงมีชีวิตชนิดใหม่ ข้อเสีย คือการทําให้เกิดโรคและ
ความผิดปกติของร่างกาย
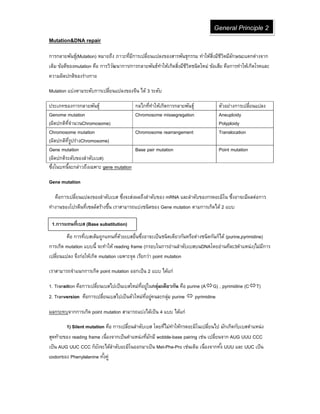
Mutation แบ่งตามระดับการเปลียนแปลงของจีน ได้ 3 ระดับ
ประเภทของการกลายพันธุ์ กลไกทีทําให้เกิดการกลายพันธุ์ ตัวอย่างการเปลียนแปลง
Genome mutation
(ผิดปกติทีจํานวนChromosome)
Chromosome missegregation Aneuploidy
Polyploidy
Chromosome mutation
(ผิดปกติทีรูปร่างChromosome)
Chromosome rearrangement Translocation
Gene mutation
(ผิดปกติระดับของลําดับเบส)
Base pair mutation Point mutation
ซึงในบทนีจะกล่าวถึงเฉพาะ gene mutation
Gene mutation
คือการเปลียนแปลงของลําดับเบส ซึงจะส่งผลถึงลําดับของ mRNA และลําดับของกรดอะมิโน ซึงอาจะมีผลต่อการ
ทํางานของโปรตีนทีเซลล์สร้างขึน เราสามารถแบ่งชนิดของ Gene mutation ตามการเกิดได้ 2 แบบ
คือ การทีเบสเดิมถูกแทนทีด้วยเบสอืนซึงอาจะเป็นชนิดเดียวกันหรือต่างชนิดกันก็ได้ (purine,pyrimidine)
การเกิด mutation แบบนี จะทําให้ reading frame (กรอบในการอ่านลําดับเบสบนDNAโดยอ่านทีละ3ตําแหน่ง)ไม่มีการ
เปลียนแปลง จึงก่อให้เกิด mutation เฉพาะจุด เรียกว่า point mutation
เราสามารถจําแนกการเกิด point mutation ออกเป็น 2 แบบ ได้แก่
1. Transition คือการเปลียนเบสไปเป็นเบสใหม่ทีอยู่ในกลุ่มเดียวกัน คือ purine (A G) , pyrimidine (C T)
2. Tranversion คือการเปลียนเบสไปเป็นตัวใหม่ทีอยู่คนละกลุ่ม purine pyrimidine
ผลกระทบจากการเกิด point mutation สามารถแบ่งได้เป็น 4 แบบ ได้แก่
1) Silent mutation คือ การเปลียนลําดับเบส โดยทีไม่ทําให้กรดอะมิโนเปลียนไป มักเกิดกับเบสตําแหน่ง
สุดท้ายของ reading frame เนืองจากเป็นตําแหน่งทีมักมี wobble-base pairing เช่น เปลียนจาก AUG UUU CCC
เป็น AUG UUC CCC ก็ยังจะได้ลําดับอะมิโนออกมาเป็น Met-Phe-Pro เช่นเดิม เนืองจากทัง UUU และ UUC เป็น
codonของ Phenylalanine ทังคู่
1.การแทนทีเบส (Base substitution)
General Principle 2
- 2. 2) sense mutation คือ การเกิด
เป็นโปรตีนปกติในร่างกายเช่นเดิม ความผิดปกติจากการเกิด
3) missense mutation คือ การเกิด
ไม่สามารถกลับมาทํางานได้เหมือนเดิม
4) nonsense mutation คือ การเกิด
โปรตีนสายสันกว่าปกติ
5) regulatory mutation คือการเกิ
การสังเคราะห์โปรตีนมากขึน หรือน้อยลง หรือสังเคราะห์ไม่ได้เลย
6) RNA processing mutation
การเปลียนลําดับเบสบริเวณทีจําเพาะต่อการตัด
คือ การทีมีเบสบนสาย DNA เพิมขึนมา หรือหายไป
ให้ reading frame บริเวณหลังต่อจุดเกิด mutation
ทังหมด เรียกว่า frameshift mutation
Regulation of gene expression
- โครงสร้างของโครโมโซม คือ Chromatin
Protein ทีชือว่า Histone (ซึงเป็นโปรตีนทีมีประจุบวก ทําให้ยึดเกาะกับ
ได้ดี) เกาะกันเป็น Nucleosome และพันกันเป็น
2. การเพิม/ขาดหายของลําดับเบส
คือ การเกิด point mutation แล้วทําให้กรดอะมิโนเปลียนไป แต่ยังสามารถทํางานได้
เป็นโปรตีนปกติในร่างกายเช่นเดิม ความผิดปกติจากการเกิด sense mutation จึงน้อยหรือแทบไม่มี
คือ การเกิด point mutation ทีทําให้กรดอะมิโนของโปรตีนเปลียนไป แล้วโปรตีน
คือ การเกิด point mutation ทีทําให้เกิด stop codon(UAA,UAG,UGA
คือการเกิด point mutation ของgene ทีบริเวณ promoter และ Enhancer
การสังเคราะห์โปรตีนมากขึน หรือน้อยลง หรือสังเคราะห์ไม่ได้เลย
6) RNA processing mutation คือ การเกิด point mutation ทีส่งผลต่อกระบวนการตัดแต่ง
การตัด RNA (RNA splicing) ทําให้ได้สาย mRNA ทีผิดปกติ
เพิมขึนมา หรือหายไป 1 ตัวหรือมากกว่า การเกิด mutation ตามกลไกนีจะทํา
mutation เลือนไปทังหมด จึงเกิดกรดอะมิโนหลังตําแหน่ง mutation
Chromatin ซึงประกอบด้วย DNA กับ
ซึงเป็นโปรตีนทีมีประจุบวก ทําให้ยึดเกาะกับDNA
และพันกันเป็น Chromatin
ขาดหายของลําดับเบส (Base insertion/deletion)
แล้วทําให้กรดอะมิโนเปลียนไป แต่ยังสามารถทํางานได้
ทีทําให้กรดอะมิโนของโปรตีนเปลียนไป แล้วโปรตีน
UAA,UAG,UGA) จึงได้
Enhancer ทําให้มี
ทีส่งผลต่อกระบวนการตัดแต่ง mRNA เช่น
ตามกลไกนีจะทํา
mutation ผิดไป
- 3. - ขนาดของโครโมโซมนันสําคัญหรือไม่ ?
มากกว่าเรา แต่เรามีวิวัฒนาการสูงกว่าพืช
- จีนของยูคาริโอตนันจะอยู่กันเป็นส่วนๆ ได้แก่
คือ ส่วนทีเราใช้
เราพบว่า
- เรา
code
เป้าหมาย
- ในการควบคุมการแสดงออกเราต้องรู้จัก
1. Promoter
บนupstreamของ DNA ซึงบริเวณนันมีรหัส
gene transcription ลดลง
2. Enhancer: บริเวณบน DNA ทีจับกับTranscription factor
3. Silencer: บริเวณทีตัว repressor มาจับ
การควบคุมการแสดงออกของจีนยูคาริโอตแบ่งเป็น
1.การควบคุมระดับจีน
1.1 การเพิมจํานวนจีน (Gene Amplification)
1.2 การจัดเรียงตัวใหม่ของDNA (DNA Rearrangement)
1.3 การเติมหมู่เมทิลของ DNA
บริเวณทีมีเบส CpG จํานวนมากอยู่ตอนต้นของจีนโครงสร้างทีเรียกว่า
แสดงออก โดยเมทิลจะไปยับยังการจับของ
คําตอบคือ ไม่เชิง เพราะพืชมีโครโมโซมใหญ่กว่าเรา และสร้างสารได้
มากกว่าเรา แต่เรามีวิวัฒนาการสูงกว่าพืช
จีนของยูคาริโอตนันจะอยู่กันเป็นส่วนๆ ได้แก่ Exons กับ Introns
คือ ส่วนทีเราใช้codeเป็น Protein, RNA แต่ส่วนทีเหลือเป็น intro
เราพบว่า Introns บางตัวมาควบคุมการแสดงออกของจีน
เราจําต้องควบคุมการแสดงออกของจีน เพราะ การแสดงออกของจีน
code RNA หรือ Biologically Active Protein ออกมาเพือแสดงออํา
เป้าหมาย และเหมาะสม เพราะร่างกายทีควบคุมความผิดปกติไม่ได้ก็จะเ
ในการควบคุมการแสดงออกเราต้องรู้จัก 3 ตัวนี
Promoter: บริเวณที RNA polymerase และ Transcription factor
ซึงบริเวณนันมีรหัสATมาก จึงเรียกว่า TATA Box ถ้า Promoter ผิดปกติ เราจะพบว่าจํานวน
Transcription factor แล้วเพิมระดับการถอดรหัสของจีน
มาจับ
การควบคุมการแสดงออกของจีนยูคาริโอตแบ่งเป็น 3 ระดับ คือ
(Gene Amplification)
DNA (DNA Rearrangement)
DNA (DNA Methylation) คือ การเติมหมู่เมทิลให้กับ Cytosine(C)
จํานวนมากอยู่ตอนต้นของจีนโครงสร้างทีเรียกว่า CpG island ซึงจะทําให้จีนนันไม่ทํางานหรือ ไม่
ลจะไปยับยังการจับของTranscription factor
DNA Rearrangemen
*Enhancer และ Silencer จะอยู่ใกล้กัน ไกลกัน หรืออยู่ในจีนทีมันควบคุมก็ได้
คําตอบคือ ไม่เชิง เพราะพืชมีโครโมโซมใหญ่กว่าเรา และสร้างสารได้
Introns ซึง Exons
introns ซึงปัจจุบัน
การแสดงออกของจีนจะต้อง
แสดงออําให้ได้ตาม
ร่างกายทีควบคุมความผิดปกติไม่ได้ก็จะเกิดโรค
Transcription factor มาเกาะ
เราจะพบว่าจํานวน
Cytosine(C) ทีC5
ซึงจะทําให้จีนนันไม่ทํางานหรือ ไม่
DNA Rearrangement
จะอยู่ใกล้กัน ไกลกัน หรืออยู่ในจีนทีมันควบคุมก็ได้
- 4. 2.การควบคุมระดับการถอดรหัส
โดยปกติเราจะมีการถอดรหัสในระดับตํา เพราะเซลล์เก็บ DNA ไว้ในรูป Nucleosome ซึงปิด Promoter ไว้ไม่
ถอดรหัส ดังนันการถอดรหัสจังต้องเป็นแบบ Positive Regulation คือ ต้องมีตัวกระตุ้นจึงจะเกิดการถอดรหัส เช่น
Transcription factor protein (TAP), ตัวเพิมประสิทธิภาพ (Enhancer)
2.1 การควบคุมการถอดรหัสโดยโครงรูปโครมาติน
โครมาตินจะถูกเก็บไว้ 2 แบบ คือ
Heterochromatin Chromatin จะอยู่กันอย่างหนาแน่น บริเวณนีมีการถอดรหัส
Euchromatin บริเวณนีอยู่กันอย่างหลวมๆ โดยอาจพบNucleosomeน้อย
หรือไม่พบเลย ซึงบริเวณนีมีการถอดรหัสสูง
นอกจากนียังมีการดัดแปลง Histone (HIstone
Modification) จะมีผลต่อการถอดรหัสด้วย เช่น
(1) Histone Acetylation คือ การเติมหมู่
Acetyl ให้กับHistone โดยใช้เอนไซม์ Histone
acetyl transferase (HAT) เพือทําให้ Histone
กลายเป็นAcetylhistone ซึงปกติ Histone มีประจุ
บวกและจับกับ DNA แต่ Acetylhistone เป็น
Neutral และไม่จับกับ DNA (เพราะ DNA มีประจุ
ลบจากฟอสเฟต) ทําให้เกิดการแสดงออกได้
หลังจากเราแสดงออกแล้ว Acetylhistone จะถูก
เปลียนกลับมาเป็น Histone โดยใช้เอนไซม์
Histone deacetylase
(2) Histone Methylation จะทําให้Histoneรวมตัวกันหนาแน่นมากขึน ทําให้กระบวนการถอดรหัสเกิดขึนได้ยาก
2.2 การควบคุมโดยโปรตีนกระตุ้นกระบวนการถอดรหัส(Transcriptional Activator Protein:TAP)
โปรตีนนีจะทําหน้าทีกระตุ้นกระบวนการสังเคราะห์ mRNA (DNA Transcription) จะมีองค์ประกอบสําคัญ 2
ส่วน คือ ส่วนทีจับกับสายDNA (DNA binding domain) กับส่วนทีกระตุ้นกระบวนการสังเคราะห์RNA โดยโปรตีนTAP
จะช่วยให้RNA polymeraseจับกับPromoterได้ดีขึน และทําให้DNAคลายตัวจากHistone ทําให้transcription complex
สามารถเข้ามาจับกับPromoterได้ โดยเราสามารถจําแนกTAPตัวคุณสมบัติและลําดับของกรดอะมิโนของส่วนทีจับกับ
ดีเอ็นเอได้หลายชนิด เช่น Helix-turn-helix, Zinc finger และ Leucin zipper
- 5. ตัวอย่าง TAP
3.การควบคุมระดับแปลรหัส
การควบคุมในระดับนีเป็นการควบคุมทีให้ผลเร็วกว่าการควบคุมในระดับอืนๆ โดยมีอายุเฉลียและความ
เสถียรของ mRNAเป็นตัวควบคุมอัตราเร็วและปริมาณการสังเคราะห์โปรตีน
การสังเคราะห์ดีเอ็นเอ (DNA replication)
- เกิดที nucleus มีเอนไซม์ทีใช้ในการสังเคราะห์ คือ
- โดยเริมทีตําแหน่งจําเพาะทีเรียกว่า Origin of replication
เกลียวออกได้ในสองทิศทาง (bidirect
- จะมี single-strand DNA-binding protein
กันอีก
- และะมี enz DNA gyrase (DNA topoisomerase II)
การควบคุมในระดับนีเป็นการควบคุมทีให้ผลเร็วกว่าการควบคุมในระดับอืนๆ โดยมีอายุเฉลียและความ
เป็นตัวควบคุมอัตราเร็วและปริมาณการสังเคราะห์โปรตีน
(DNA replication)
มีเอนไซม์ทีใช้ในการสังเคราะห์ คือ DNA polymerase
Origin of replication โดยจะมี enz helicase เข้ามาแยกสายดีเอ็นเอให้คลาย
(bidirection) ของจุดเริมต้น โดยมีจุดแยกสายทีเรียกว่า replicaton fork
binding protein (SSB) มาเกาะกับดีเอ็นเอสายเดียวทังสองสาย เพือไม่ให้กลับมาเข้าคู่
enz DNA gyrase (DNA topoisomerase II) ช่วยให้ดีเอ็นเอเกลียวคู่ทีพันกันแน่นคลายตัวออก
การควบคุมในระดับนีเป็นการควบคุมทีให้ผลเร็วกว่าการควบคุมในระดับอืนๆ โดยมีอายุเฉลียและความ
เข้ามาแยกสายดีเอ็นเอให้คลาย
replicaton fork
มาเกาะกับดีเอ็นเอสายเดียวทังสองสาย เพือไม่ให้กลับมาเข้าคู่
นคลายตัวออก
- 6. - จากนัน DNA สายเดียวจะมี enz primase สร้าง RNA primer มาเกาะ เพือเป็นจุดเริมต้นให้ DNA polymerase III
สังเคราะห์ DNA โดยจะเคลือนไปในทิศ 3’ ไป 5’ ของสาย template (5’ ไป 3’ ของสายใหม่) จะได้ทัง leading
strand และ lagging strand (Okazaki fragment)
- เมือสร้างสาย DNA มาถึง RNA primer ทีอยู่ข้างหน้า DNA polymerase III จะแยกออกเพือให้ DNA polymerase
I มาจับแทนเพือตัด RNA primer และสร้าง DNA แทนที RNA primer ก่อนทีจะแยกตัวออกแล้วให้ DNA ligase
มาเชือมต่อสาย DNA
การสังเคราะห์อาร์เอ็นเอ (RNA transcription)
- เกิดที nucleus การสังเคราะห์ RNA จะใช้ DNA เป็นแม่แบบและใช้ Enz RNA polymerase สังเคราะห์ RNA ใน
ทิศ 5’ ไป 3’ โดย RNA polymerase จะไม่มี Proofreading activity เหมือน DNA polymerase
- ต้องการ DNA เพียงสายเดียวเป็นแม่แบบ เรียกสายทีเป็นแม่แบบว่า template strand ส่วนอีกสายเรียกว่า non-
template strand หรือ coding strand เพราะมีการเรียงลําดับเบสเหมือน mRNA ทีสังเคราะห์ขึนมา
- การเข้าคู่กันของ template strand และ RNA nucleotide A-U,T-A,C-G,G-C
ตัวอย่าง DNA template 5’-TACGGCAATCT-3’
จะได้ mRNA 3’-AUGCCGUUAGA-5’
- หลังจากได้ mRNA แล้วจะต้องผ่านการตัดแต่ง (post-transcriptional RNA processing มีอยู่ 3 ขันตอน คือ
1.การเติม 5’cap คือการเติม 7-methylguanosine
เข้าทีปลาย 5’ ของ mRNA
2.การเติม 3’tail ทีปลาย 3’ ของ mRNA
ประกอบด้วย poly A tail (มี enz
riboendonuclease ตัด mRNA บริเวณทีจะเติม
poly A tail (มี AAUAAA เป็น cleavage signal)
แล้วเติม poly A เข้าไป เชือมด้วย enz
polyadenylate polymerase)
3.RNA Splicing ปกติ mRNA ทีถอดรหัสมาจะ
ปกด.ส่วนทีถูกแปลรหัสเป็นโปรตีน เรียกว่า Exon และส่วนทีไม่ถูกแปลรหัสเป็นโปรตีน เรียกว่า intron โดย
กระบวนการ splicing จะตัด intron ออก แล้วนํา exon มาเชือมต่อกัน จะได้ Mature mRNA
เพิมเติม RNA polymerase I สร้าง rRNA
RNA polymerase II สร้าง mRNA ถูกยับยังโดย alpha-amanitin พบในเห็ด amanita ทําให้เกิด liver failure ได้
RNA polymerase III สร้าง tRNA
- 7. การสังเคราะห์โปรตีน (Protein translation)
- เกิดใน cytoplasm โดย ribosome จะแปลรหัสลําดับเบสบน mRNA
จาก 5’ ไป 3’ แล้วสังเคราะห์โปรตีนจาก N-terminal ไป C-terminal
- โดยจะอ่านรหัสบน mRNA ทีละ 3 รหัส เรียกว่า codon
Start codon AUG (Met)
Stop codon UAA UAG UGA
** ทุก codon จะให้กรดอะมิโนได้หมด ยกเว้น stop codon
** ควรรู้... tryptophan (UGG) และ methionine (AUG)
- tRNA จะจับกับ codon โดยมีเบสตรงข้ามกับ codon เรียก
anticodon เป็นตัวกระตุ้นและนําพากรดอะมิโนชนิดต่างๆ
- มี ribosome เป็นตัวทําหน้าทีสร้าง peptide bond และรองรับ
mRNA และ tRNA
กระบวนการสังเคราะห์โปรตีน มี 3 ขันตอน
1. Initiation - initiation factors นํา 30s ribosome มาจับกับ mRNA บริเวณ Shine-Dalgarno sequence
- start codon (AUG) จะอยู่ที P-site ของไรโบโซม และมี fMet-tRNA มาเกาะที start codon โดยนํา Methionine
ติดไม้ติดมือมาด้วย
- 50s ribosome มาจับกับ 30s ribosome& fMet-RNA กลายเป็น 70s initiation complex
2. Elongation – จะมี tRNA ตัวทีสองเข้ามาที A-site ของไรโบโซม แล้วมีการสร้าง peptide bond เชือมระหว่าง
กรดอะมิโนที P-site และ A-site โดย enzyme peptidyltransferase
3. Termination – จะสินสุดเมือถึง stop codon (UAA UAG UGA)
- 8. The genetic code
รหัสพันธุกรรมใช้สือความหมายระหว่างลําดับเบสใน mRNA กับลําดับกรดอะมิโนในสารพอริเพปไทด์ โดยใช้
รหัสสามชุดหรือ codon โดยพบว่ากรดอะมิโน 1 ตัวต้องใช้เบส 3 หน่วยทีเรียงติดกันเป็นตัวกําหนด มีรหัส 64 รหัสเป็น
รหัสของกรดอะมิโน 20 ชนิด ดังนันกรดอะมิโน 1 ชนิดอาจมีรหัสมากกว่า 1 รหัส โดยใน 64 รหัสเป็น start codon 1 รหัส
คือ AUG ซึงเป็นรหัสของกรดอะมิโน Met มีรหัสหยุด 3 รหัสคือ UAA, UAG และ UGA ซึงไม่ได้เป็นรหัสสําหรับกรดอะมิ
โนชนิดใดเลย โดยมีกรดอะมิโนชนิดทีมีรหัสเพียงรหัสเดียวคือ methionine กับ tryptophan ในกรณีทีกรดอะมิโนชนิดนันๆ
มีรหัสมากกว่า 1 รหัส จะสังเกตได้ว่า รหัสเหล่านีส่วนใหญ่มีความแตกต่างต่างกันทีเบสตัวที 3 ดังนันเบส 2 ตัวแรกของ
รหัสจะเป็นตัวบ่งบอกความจําเพาะว่าเป็นกรดอะมิโนชนิดใด
เดิมเชือว่ารหัสพันธุกรรมเป็นรหัสสากล ไม่ว่าจะเป็น Eukaryote หรือ Prokaryote ก็จะสือความหมายถึงกรดอะมิ
โนชนิดเดียวกัน แต่ต่อมาพบว่ารหัสบางตัวในไมโตคอนเดรียสือความหมายต่างจากพันธุกรรมสากล
Structure & Function of tRNA
tRNA เป็น RNA ทีมีขนาดเล็กทีสุด ประกอบด้วย RNA
เพียง 1 สาย แต่บางส่วนเป็นองค์ประกอบของกันและกันทํา
ให้เกิดเป็นเกลียวคู่ขึนในบางส่วนของโมเลกุล และทําให้ส่วน
ทีเหลือเปิดเป็นวงขึน และมองดูคล้ายใบClover โดยวงที
เกิดขึนมี 4 วง ประกอบด้วย DHU loop, anticodon loop,
extra or variable loop และpseudouridine loop
tRNA ทําหน้าทีเป็นตัวพากรดอะมิโนมายังไรโบโซม
เพือประกอบกันเป็นสารพอลิเปปไทด์ได้อย่างถูกต้อง tRNA
แต่ละชนิดจะมีความจําเพาะต่อชนิดของกรดอะมิโนคือ
สามารถรับและพากรดอะมิโนได้เพียงชนิดเดียวเท่านัน
Structure & Function of Ribosome
Ribosome เป็นออร์กาแนล ทีประกอบด้วย rRNA 65% และโปรตีนชนิดต่างๆ 35% โดยส่วนของโปรตีนทํา
หน้าทีเกียวกับโครงสร้างและเป็นเอนไซม์ได้ ไรโบโซมของโปรคาริโอตและยูคาริโอตมีขนาดไม่เท่ากัน โดยribosome
ของยูคาริโอตมีค่าสัมประสิทธิการตกตะกอนเท่ากับ 80S ประกอบด้วยหน่วยย่อย 40S และ 60S ส่วนไรโบโซมของโปร
คาริโอตมีค่าสัมประสิทธิการตกตะกอนเท่ากับ 70S ประกอบด้วยหน่วยย่อย 30S และ 50S พบไรโบโซมอยู่ทังไซโท
ซอล ไมโทคอนเดรีย และในเอนโดพลาสมิกเรติคูลัม โดยเชือว่าไรโบโซมทีอยู่ทีเอนโดพลาสมิกเรติคูลัม โดยเชือว่า
ไรโบโซมทีอยู่ทีเอนโดพลาสมิกเรติคูลัมนีเป็นแหล่งผลิตโปรตีนเพือส่งออกไปใช้ทีอืนภายนอกเซลล์ ส่วนพวกทีอยู่ในไซ
โมซอลนัน ทําหน้าทีผลิตโปรตีนทีใช้ภายในเซลล์เอง
- 10. Translation mRNA>>>Protein
- mRNA
- tRNA + amino acid (aminoacyl-tRNA)
- rRNA + ribosome
- Factor, Elongation factors (Tu,Ts ) , Initiation factors (IF I, IF II, IF III )
Processing
IF-3 + 30S ไปจับ Shine-Dalgano sequence AUG อยู่ตรง P site
ใน Euc. ไม่มี fMet นํา fMet-tRNA มาที P site
30S initiation complex
70S initiation complex
Tu-GTP พา Aa-tRNA ไปจับที A site
สลาย GTP
Tu หลุดออกจากไรโบโซม
Tu + Ts
Tu + Ts Tu-GTP + Ts
สร้างพันธะ peptide
Translation *ribosome เลือน A site ว่าง เลือนไปถึง Stop codon
RF I หรือ RF II + RF III-GTP จับกับ stop codon
สาย Polypeptide หลุดออกจาก tRNA ที P site Polypeptide เป็นอิสระ
RF, +RNA, mRNA แยกตัวจาก ribosome
30S กับ 50S แยกกัน
+ 50S
ระยะเริมต้น
Eukaryote ใช้Factor 9 ตัว
ระยะเริมยาว
การสังเคราะห์ Aa-tRNA
Amino acid + Enzyme + ATP
Enzyme-(Aminoacyl – AMP) + PPi + tRNA
Aminoacyl-tRNA + AMP + Enzyme
Enzyme = aminoacyl-tRNA synthetase
RF-1 + UAA, UAG
RF-2 + UAA, UGA
RF-3 + GTP
ระยะสุดท้าย
สรุป ใช้1ATP+2GTP / 1Aa
AMP
2Pi
- 11. Regulation of translation
Prokaryote
1. การควบคุมโปรตีนทีสังเคราะห์จาก Polycistronic mRNA
ในโปรคาริโอต mRNA 1 สาย สามารถสังเคราะห์ออกมาได้โปรตีนหลายชนิด และการสังเคราะห์ก็อาจเกิดขึน
พร้อมกันหลายชุด ซึงทําให้โปรตีนแต่ละชนิดมีปริมาณเท่ากัน แต่ในความเป็นจริงกลับพบแตกต่างกันมาก โดย
mRNA ของโปรตีนทีอยู่ในลําดับต้นๆจะถูกแปลเป็นโปรตีนมากกว่าทีอยู่ส่วนปลาย โดยขึนอยู่กับแรงยึดเหนียว
ระหว่างไรโบโซม mRNA และระยะห่างระหว่างรหัสหยุดของ mRNA รหัสแรก กับรหัสเริมของ mRNAรหัสถัดไป
พบว่า ไรโบโซมส่วนใหญ่มักหลุดออกจากสาย mRNA เมือถึงรหัสหยุดของโปรตีนตัวแรก
2. การควบคุมการสังเคราะห์โปรตีนแบบ Transitional grameshift
ในการสังเคราะห์โปรตีนบางชนิด mRNA ของโปรตีนนันมีรหัสหยุดอยู่ในสายการสร้าง ถ้าในเซลล์มีโปรตีนชนิดนัน
มากก็จะหยุดการสังเคราะห์ทีรหัสหยุดนัน ทําให้โปรตีนทีสร้างออกมาไม่สมบูรณ์ แต่ถ้าโปรตีนนันขาดแคลนจะเกิด
การอ่านข้ามเบสไป 1 ตําแหน่ง ทําให้การแปลรหัสในตําแหน่งถัดไปเคลือนไปจากเดิม เรียกว่าเกิด transitional
freameshift พบมากใน retrovirus
3. การควบคุมการสังเคราะห์โปรตีนโดย Transitional repressor
เป็นการควบคุมการสังเคราะห์โปรตีน โดยโปรตีนตัวทีผลิตเป็นการควบคุมโดยการยับยังแบบย้อนกลับ โดยเกิด
ขึนกับ ribosomal protein โปรตีน Elongation factor EF-G และ EF-Tu เป็นต้น
4. การควบคุมการสังเคราะห์โปรตีนโดย Antisense RNA
Antisense RNA เป็นสายRNA สันๆทีสามารถจับจําเพาะกับ mRNA ทีปลาย 5’ ทําให้ขัดขวางการสังเคราะห์
โปรตีนชนิดนัน เช่น ควบคุมการสังเคราะห์โปรตีน CAP ใน lac operon
- 12. Eukaryote
1. การควบคุมอายุเฉลียและความเสถียรของ mRNA
อายุเฉลียของ mRNA เป็นปัจจัยหนึงในการควบคุมปริมาณและอัตราเร็วในการผลิตโปนตีน ตัวอย่างเช่น การสร้าง
โปรตีนเส้นไหมของตัวไหม โดยจะสร้าง mRNA ของเส้นไหมให้มีอายุเฉลียยาวกว่าปกติ ช่วยลดเวลาในการ
สังเคราะห์เส้นไหม หรือการสังเคราะห์โปรตีนเคซีน (Casine) ในหญิงให้นมบุตร โดยฮอร์โมนโพรแลกติน จะช่วย
ให้ mRNA ของเคซีนมีความเสถียรมากขึน
2. การควบคุมการแปลรหัสโดยการสังเคราะห์เป็นพอลิโปรตีน
ในยูคาริโอตจะมีการสังเคราะห์โปรตีนสายยาวทีเรียกว่า Polyprotein เป็นโปรตีนเริมต้น จากนันจึงมีกระบวนการ
ตัดสายโปรตีนนีอย่างจําเพาะ ทําให้ได้โปรตีนหลายๆชนิด การผลิตโปรตีนโดยวิธีนี เป็นการควบคุมให้โปรตีน
ผลผลิตทีเกิดจากการตัดสายเริมต้นนีมีปริมาณเท่ากัน หรือใกล้เคียงกัน
3. การควบคุมอัตรารวมของกระบวนการแปลรหัส
เป็นกลไกควบคุมการผลิตโปรตีนโดยใช้สารทีจะมารวมตัวกันเป็นตัวควบคุม เช่น การสังเคราะห์โปรตีนโกลบิน
โดยจะมีฮีมเป็นตัวควบคุมการสังเคราะห์ ถ้ามีฮีมในเซลล์มาก ฮีมจะยับยังกระบวนการสังเคราะห์ฮีม และกระตุ้น
ให้มีการสังเคราะห์โปรตีนโปรตีนโกลบินเพิมขึน เป็นต้น
4. การระงับการทํางานของ mRNA โดย RNA interference
RNA interference เป็นกระบวนการทีชิน RNA ขนาดเล็กไปจับกับปลาย 3’ UTR ของ mRNA และยับยังการ
สังเคราะห์โปรตีน หรือทําให้เกิดการทําลายสายmRNA
Post – translational Modification
- จัดรูปร่าง
- Inactive active
- เติม Saccharides
- เติม Substitute groups ex acetyl, phosphate, methyl, carboxyl
- หลังและถูกกระตุ้นให้ทํางาน
- สร้าง disulfide cross-links
- 13. การเติมโมเลกุลของคาร์โบไฮเดรต (glycosylation
ไกลโคโปรตีน (glycoprotein) เกิดจากการเติมสายคาร์โบไฮเดรตลงบนโมเลกุลของโปรตีน กรดอะมิโนในโปรตีน
ทีเป็นตัวเชือมต่อกับสายคาร์โบไฮเดรต ได้แก่ แอสพาราจีน
สายคาร์โบไฮเดรตนีต้องอาศัยการทํางานของกลุ่มเอนไซม์
ใหญ่ของคาร์โบไฮเดรตทีถูกนํามาเติม ได้แก่ แมนโนส
(xylose) N-acetylglucosamine (NAcGlc) และ N-acetylneuraminic acid (N
glycosylation)
เกิดจากการเติมสายคาร์โบไฮเดรตลงบนโมเลกุลของโปรตีน กรดอะมิโนในโปรตีน
ทีเป็นตัวเชือมต่อกับสายคาร์โบไฮเดรต ได้แก่ แอสพาราจีน (asparagine) เซอรีน (serine) และทรีโอนีน (threonine
าศัยการทํางานของกลุ่มเอนไซม์ glycosyltransferase ซึงพบอยู่ในเอนโดพลาสมิกเรติคูลัม ส่วน
ใหญ่ของคาร์โบไฮเดรตทีถูกนํามาเติม ได้แก่ แมนโนส (mannose) กลูโคส (glucose) กาแลกโทส (galactose
acetylneuraminic acid (NAN) หรือ กรดเซียริก (sialic acid) เป็นต้น
เกิดจากการเติมสายคาร์โบไฮเดรตลงบนโมเลกุลของโปรตีน กรดอะมิโนในโปรตีน
threonine) การเติม
ซึงพบอยู่ในเอนโดพลาสมิกเรติคูลัม ส่วน
galactose) ไซโลส
- 14. การเติมหมู่ฟอสเฟต (phosphorylation)
ปฏิกิริยาการเติมหมู่ฟอสเฟตให้กับกรดอะมิโนบางตัวในสายพอลิเพปไทด์โดยเอนไซม์ protein kinase เป็น
กระบวนการหนึงทีเซลล์ใช้ควบคุมหรือกําหนดการทํางานของโปรตีน กรดอะมิโนมักจะถูกเติมหมู่ฟอสเฟต ได้แก่ เซอ
รีน (serine) ทรีโอนิน (threonine) และไทโรซีน (tyrosine)
Ex Insulin
Translation folding, oxidation, Signal peptide cleavage vesicle packaging ตัด C-Chain
Ribosome
- Cytosol >>> โปรตีนทีใช้ในเซลล์
- ER ER Lumen >> exocytosis
ER membrane >> Protein membrane
Inhibition protein synthesis
- Puromycin : ไปจับกับ A site คล้าย A-tRNA
- Tetracycline : จับกับ 30S ป้องกันไม่ให้ tRNA ไปเกาะ A site
- Erythromycin : จับกับ 50S >>> translocation (เคลือนที) ไม่ได้
- Chloramphenicol: สร้าง Peptide-bond ไม่ได้ใน bacteria mitochondria chloroplast
block peptidyl translation
- Cycloheximide: Block peptidyl transfer of 50S
- Streptomycin: ไม่ให้ fMet-tRMA จับ 30S และทําให้อ่านผิดด้วย
- Diptheria toxin: ยับยัง IF II ใน Eukaryote
- Ricin: จาก caster benan : ทําให้ 60s ไม่ทํางาน
สร้าง disulfide
Mature insulin
- 15. Protein degradation
protein degradetion เป็นกระบวนการย่อยสลายโปรตีนภายในเซลล์ อาจเป็นโปรตีนจากกระบวนการ
หรือโปรตีนทีไม่ได้ช้งานภายในเซลล์ แบ่งออกเป็น
organelle และCytosolic อาศัยproteasomes, multienzyme complexes
-
-
เป็นกระบวนการย่อยสลายโปรตีนภายในเซลล์ อาจเป็นโปรตีนจากกระบวนการ
หรือโปรตีนทีไม่ได้ช้งานภายในเซลล์ แบ่งออกเป็น 2 วิธี คือ LYSOSOMAL อาศัยเอมไซม์ Proteases
proteasomes, multienzyme complexes ใน cytoplasm
เป็นกระบวนการย่อยสลายโปรตีนภายในเซลล์ อาจเป็นโปรตีนจากกระบวนการendocytosis
Proteasesใน acidic
- 16. การหาลําดับนิวคลีโอไทด์ (DNA sequencing)
หรือการหาลําดับเบสของ DNA ซึงประกอบด้วย A G C T อาศัยปฏิกิริยา
และเทคนิคต่างๆมากมาย ซึงเมือเราทราบลําดับเบสของ DNA ในสิงมีชีวิตต่างๆ จะ
ทําให้เราเข้าใจพันธุกรรมของสิงมีชีวิตนันๆ และสามารถนําความรู้หรือลําดับเบสที
ได้มาใช้หลายๆสาขา เช่น วินิจฉัยโรค,เทคโนโลยีชีวภาพ และนิติเวชศาสตร์ เป็นต้น
ปัจจุบันหลักการทีใช้ในการทํา DNA Sequencing ประยุกต์มาจากขันตอนวิธี
ของ Frederick Sanger นักวิทยาศาสตร์ชาวอังกฤษ จาก มหาวิทยาลัยแคมบริดจ์ โดยมีหลักการดังนี
1. DNA Sequencing Reaction คล้ายกับการเริมต้นปฏิกิริยา PCR เพือเพิมจํานวน DNA โดยให้ความร้อนเพือให้
DNA แยกเป็นสายเดียว จากนัน Primer ไปเกาะเพือเกิด elongation ของ DNA ต่อไป
โดยใน substrate จะประกอบด้วย
(1) DNA Template – ซึงเป็นสายคู่
(2) free nucleotides – dATP,dGTP,dCTP,dTTP
(3) DNA Primer ขนาดสันๆ
(4) enzyme Taq polymerase เพือเร่งปฏิกิริยา polymerization
(5) dideoxynucleotides(ddNTs) คือ NTs ทีไม่มีหมู่
Hydroxyl ทีตําแหน่ง 3’ (ddATP, ddGTP, ddCTP, ddTTP)
เป็นส่วนสําคัญในปฏิกิริยาเพราะทําให้เกิด Chain termination
2. ทุกๆ cycle ในการทําปฏิกิริยาเมือมี ddNTs ใน substrate จะมีโอกาสที ddNTs
เข้าไปทําปฏิกิริยา elongation แต่จะทําให้ Taq polymerase หยุดทําปฏิกิริยา
ทันทีและสินสุดด้วย ddNTs นัน เรียกว่าวิธีการนีว่า Chain Termination
Methods
3. เราจะพบว่า ddNTs ทีเข้าไปทําให้เกิด Chain Termination ได้ในทุกๆ
จํานวนเบสของ DNA Template นัน ทําให้ขนาดของชินส่วนหลังปฏิกิริยา
มีหลากหลายตามจํานวนเบสของ ชินส่วนนันๆ โดยตรวจสอบจากการทํา
Gel Electrophoresis พบว่าชินส่วนทีอยู่ไกลสุดจากจุดเริมจะมีจํานวนเบสน้อย
สุด และตรงกันข้าม (ในแต่ละ lane จะมี ddNTs เป็น terminator ต่างกัน)
- 17. 4.
โดย
Electrophoresis
ได้โดยใช้การอ่านเอง หรือใช้เครือง
5.
(
Termination Method
ใส่
ไฟฟ้า จากนันจึงใช้เลเซอร์อ่านจากล่างขึนบนออกมาเป็นลําดับนิวคลีโอไทด์
ซึงสามารถทําได้ทีละหลายๆ ตัวอย่าง
หากจะหาลําดับเบสของจีโนมมนุษย์ซึงมีมากกว่า
และทรัพยากรมาก ซึงก็มีคนทําอยู่สองกลุ่มในการค้นหาลําดับเบสบนโครโมโซม
1. Human Genome Project เกิดจากความร่วมมือระหว่างรัฐบาลสหรัฐและห
BAC(Bacterial Artificial Chromosome)
ประมาณ 300kb แล้วใช้เทคนิค Chain termination
ผลออกมาเป็นลําดับเบสของ DNA ทังหมด
2. Celera Genomics (Craig Ventor)เป็นบริษัททุนเอกชนทีหาลําดับเบสของมนุษย์ได้สําเร็จเป็นรายแรก โดยใช้
เทคนิค Shotgun sequencing โดยนํา
10kb แล้วหาลําดับเบสของแต่ละท่อน จากนันนําท่อนทีกระจัดกระจายมาต่อกันโดยใช้คอมพิวเตอร์คํานวณและ
ให้ผลลัพธ์ออกมาเหมือน DNA ต้นแบบ ซึงทําได้ไวกว่าวิธีการข้างต้น
4. เราสามารถทําปฏิกิริยาเพือให้เกิดพร้อมๆกัน โดย ใส่ ddNTs
โดยติดฉลากเรืองแสงให้ต่างกัน เมือนํามาแยกขนาดโดย Gel
Electrophoresis แล้วเราจะอ่าน ลําดับนิวคลีโอไทด์ ได้จากล่างขึนบ
ได้โดยใช้การอ่านเอง หรือใช้เครือง Automated DNA sequencer
5. ในแลบขนาดใหญ่จะไม่ใช้ Gel แต่จะใช้วิธี Capillary electrophoresis
(Dye-terminator sequencing) แทน โดยนําตัวอย่างมาทํา Chain
Termination Method ดังกล่าวแล้วนําไป
ใส่หลอดขนาดเล็กแล้วแยกขนาดด้วย
จากล่างขึนบนออกมาเป็นลําดับนิวคลีโอไทด์
ตัวอย่างในเวลาเดียวกัน
ลําดับเบสของจีโนมมนุษย์ซึงมีมากกว่า 3,000 ล้านเบส ซับซ้อนกว่าตัวอย่างทัวๆไป จึง
คนทําอยู่สองกลุ่มในการค้นหาลําดับเบสบนโครโมโซม 23 คู่ ของคนเราดังนี
เกิดจากความร่วมมือระหว่างรัฐบาลสหรัฐและหลายประเทศ โดยสร้าง
BAC(Bacterial Artificial Chromosome) หลายๆอัน ซึงเป็น vector เก็บ genomic DNA มนุษย์อยู่ ความยา
Chain termination หาลําดับเบส แล้วจึงหาส่วนทีทับซ้อนระหว่างชิน
ทังหมด
เป็นบริษัททุนเอกชนทีหาลําดับเบสของมนุษย์ได้สําเร็จเป็นรายแรก โดยใช้
โดยนํา genomic DNA ของมนุษย์มาตัดโดยเอนไซม์โดยสุ่ม ให้มีขนาดเหลือ
แล้วหาลําดับเบสของแต่ละท่อน จากนันนําท่อนทีกระจัดกระจายมาต่อกันโดยใช้คอมพิวเตอร์คํานวณและ
ต้นแบบ ซึงทําได้ไวกว่าวิธีการข้างต้น
Shotgun sequencing
ddNTs ทัง 4 ชนิด
Gel
ล่างขึนบน ซึงทํา
Automated DNA sequencer
electrophoresis
Chain
ล้านเบส ซับซ้อนกว่าตัวอย่างทัวๆไป จึงต้องใช้ทุน
ของคนเราดังนี
ลายประเทศ โดยสร้าง Library เก็บ
มนุษย์อยู่ ความยาว
หาลําดับเบส แล้วจึงหาส่วนทีทับซ้อนระหว่างชิน เพือแปล
เป็นบริษัททุนเอกชนทีหาลําดับเบสของมนุษย์ได้สําเร็จเป็นรายแรก โดยใช้
ให้มีขนาดเหลือ 2-
แล้วหาลําดับเบสของแต่ละท่อน จากนันนําท่อนทีกระจัดกระจายมาต่อกันโดยใช้คอมพิวเตอร์คํานวณและ
- 18. ไวรัสของแบคทีเรีย (Bacteriophage)
ไวรัสของแบคทีเรียหรือ phage เป็นไวรัสทีใช้เซลล์แบคทีเรียเป็นโฮสต์ ในการเจริญเพิมจํานวน การศึกษา
เกียวกับ phage ทําให้ค้นพบเกียวกับโปรตีนและพบว่ากรดนิวคลีอิคเป็นสารพันธุกรรม (Hershey and Chase experiment)
และมีการนํา phage มาใช้กันมากในการถ่ายทอดยีนโดยวิธี transduction
รูปร่างและโครงสร้างของ phage
1. Phage ทีมีส่วนหัวและส่วนหาง
ส่วนหัว (head) มีรูปร่าง icosahedral เป็นโปรตีนทีหุ้มรอบจีโนมไว้ภายใน
ส่วนหาง (tail) มีลักษณะเป็นท่อกลวงยาว (hollow tube) หุ้มด้วยเปลือก
(sheath) ทีอาจยืดหดได้ และบางชนิดมีส่วนประกอบอืนด้วยคือ base
plate, fiber และ pin หรือ spike ความยาวของส่วนหางขึนอยู่กับชนิดของ
phage
2. Phage ทีมีรูปร่างเป็นสายยาว (filamentous phage) มีการเรียงตัวของ
capsidเป็นรูปบันไดวน (helical)
ชนิดของ phage เมือแบ่งตามลักษณะการติดเชือในแบคทีเรีย
1. Virulent phage
คือพวกทีมีการติดเชือได้แบบเดียวคือแบบทีทําให้เซลล์แบคทีเรียแตกตาย ซึงเรียกว่า lytic infection หรือ productive
infection อาจเรียก phage พวกนีว่า lytic phage, vegetative phage, virulent phage ก็ได้ วงชีวิตแบบนีเรียกว่า lytic
cycle ตัวอย่างของ phage แบบนีเช่น T4 phage
2. Temperate phage
temperate phage หมายถึง phage มีการติดเชือได้2 ลักษณะคือ
1. lytic cycle แล้วทําให้แบคทีเรียแตกสลาย
2. prophage cycle หรือ lysogenic infection ซึงไม่ทําให้เซลล์แบคทีเรียแตกสลายแต่จีโนมของ phage จะเข้าไปแทรก
อยู่ใน DNA ของแบคทีเรีย เรียกว่าเป็นกระบวนการ lysogenization
วงชีวิตของ Bacteriophage มี 2 ลักษณะ
1.Lytic pathway (Lytic-แตก)
เป็นวงจรชีวิตของ Phage ทีเมือสารพันธุกรรมจาก phage ที
เข้าไปในแบคทีเรียแล้วจะจัดตัวเองให้อยู่ในรูปคล้าย plasmid แล้ว
ดําเนินการเพิมจํานวนสารพันธุกรรม และสร้างโปรตีนต่าง ๆ ทีจาเป็น
ต่อการประกอบเป็นเซลล์ใหม่ เมือสร้างสารต่าง ๆ และประกอบตัว
เป็นอนุภาคไวรัสทีสมบูรณ์แล้วจะปล่อย Lysozyme ย่อยเซลล์เจ้าบ้าน
เพือปลดปล่อยลูกหลานไวรัสออกมา
2. Lysogenic หรือ Prophage pathway (Lyso-ซ่อนแฝง)
เป็นวงชีวิตทีเพิมจํานวนสารพันธุกรรมของ Phage ผ่านการ
แบ่งตัวของแบคทีเรีย เพราะเมือPhage ส่งสารพันธุกรรมเข้าไปใน
แบคทีเรียแล้ว สารพันธุกรรมจะปรับให้สามารถเข้าไปแทรกอยู่ใน
โครโมโซมของแบคทีเรีย(อาศัยเอนไซม์ integrase) ซึงในระหว่างนัน
- 19. สารพันธุกรรมของ Phage จะไม่กําหนดการสร้างอะไรแต่จะฝังตัวนิงๆ อยู่ในโครโมโซมของแบคทีเรีย เพิมจํานวนไป
พร้อม ๆ กับการเพิมจํานวนของแบคทีเรีย จนเมือถึง สภาวะทีเหมาะสมจึงหลุดออกจากโครโมโซมของแบคทีเรีย และ
จัดรูปแบบตัวเองเป็น plasmid ซึงทํา ให้กลับไปดํารงชีวิตในวงจร Lytic pathway ต่อไป
กระบวนการในการเกิด lysogenization
เกิดจากจีโนมของ phage มีส่วนทีเรียกว่า repressor gene ซึงเมือเข้าไปในเซลล์แบคทีเรียจะทําให้มีการ
สร้างตัวยับยังเรียกว่า repressor protein ขึนเป็นจํานวนมาก ไปเกาะกับจีโนมของแบคทีเรีย ทําให้มีผลยับยังการ
ทํางานของเอ็นไซม์ RNA polymerase ของแบคทีเรีย ทําให้ไม่เกิดการเข้าสู่ lytic cycle ถ้าตัวยับยังถูกทําลายลงเช่น
ถูกรังสี ultraviolet, X-ray สารเคมีหรือความร้อน prophage จะกลายเป็น virulent phage ทันที
bact ทีมี prophage(viral genome) แฝงอยู่จะเรียกว่า lysogen หรือ lysogenic bacteria และอยู่ในภาวะ lysogenic
lysogenic bacteria นีมีความต้านทานหรือ immunity ต่อการติดเชือซําด้วย phage ทีทําให้เกิด lysogenization นันๆ
การเพิมจํานวนของ Phage
1. การเกาะติด (Adsorption)
phage จะใช้ส่วนผิวของมันในการเกาะกับ receptor บนผิวของเซลล์แบคทีเรีย
- phage ทีมีหัวและหาง จะใช้ส่วนหาง (plate, fiber และ pin) เกาะกับผิวแบคทีเรีย
- phage ทีมีรูปร่างเป็นท่อนยาว จะใช้ส่วนปลายทังสองข้างในการเกาะกับ sex pili
ของแบคทีเรีย
- phage บางชนิดเกาะกับ flagella ของแบคทีเรียก่อนทีมันจะเลือนตัวเองลงมาเกาะ
กับผิวเซลล์โดยอาศัยการเคลือนไหวของ flagella receptor บนเซลล์แบคทีเรียอาจเป็น ผนังเซลล์ แคฟซูล(Vi antigen)
หรือ sex pili ก็ได้แล้วแต่ชนิดของ phage ในขันตอนการเกาะติดของ phage มีความจําเพาะสูงมาก ถ้าแบคทีเรียมีการ
ผ่าเหล่า เปลียนแปลงโครงสร้างของ receptor ไป phage จะไม่สามารถเกาะติดได้
2. การเข้าสู่เซลล์ (Penetration)
- phage ทีมีหางและ sheath จะหด sheath แทงท่อ (tube) ผ่านผนังเซลล์ แล้วฉีดจีโนมผ่านท่อเข้าไปในเซลล์
แบคทีเรีย ในกรณีทีมี phage จํานวนมาก อาจทําให้เกิดรูพรุนบนผนังเซลล์แบคทีเรีย และเซลล์แตกตายก่อนทีไวรัสจะ
เพิมจํานวน เรียกว่า “lysis from without”
- พวก phage ทีมีรูปร่างเป็นสายยาวจะผ่านผนังเซลล์เข้าไปทังอนุภาค และฉีดจีโนมเข้าไปในไซโตพลาสซึมของ
แบคทีเรียและทิงCapsidไว้ที cell membrane เป็นส่วนใหญ่
virulent phage จะเข้าสู่ขันตอนการเพิมจํานวนทันทีทีฉีด DNA เข้าสู่เซลล์แบคทีเรีย โดย Phage จะห้ามการสร้าง
โปรตีนทีเป็นของเซลล์ และจะย่อยทําลาย DNA ของแบคทีเรียเพือใช้เป็นสารตังต้นของการสร้างจีโนมของไวรัส
3. การ transcription,translation และ การสังเคราะห์จีโนมของไวรัส (genome replication)
มีขันตอนแตกต่างกันไปตามแต่ชนิดจีโนมของ phage ว่าเป็น DNA หรือ RNA และเป็นสายคู่หรือสายเดียว
4. การประกอบตัว (assembly)
เมือสร้างส่วนประกอบคือ จีโนมและโปรตีนต่างๆเช่น Capsid และหางแล้ว ก็จะมารวมตัวกันเป็น Phage ทีสมบูรณ์
การรวมตัวนีเกิดขึนเองโดยอัตโนมัติ เป็นขันตอน phage ทีมีหัวและหางจะเริมประกอบส่วนหัวและส่วนหางแยกกัน
และบรรจุจีโนมเข้าไปในส่วนหัวแล้วส่วนหางจึงมาประกอบเข้าไป ส่วน phage ทีเป็นสายยาว จีโนมและCapsid จะ
ประกอบตัวกันในขันตอนเดียว
- 20. 5. การออกจากเซลล์ (Release)
Phage ส่วนมากจะใช้เอ็นไซม์ lysozyme หรือเอ็นซัยม์ holin ในการย่อยผนังเซลล์แบคทีเรีย ทําให้เซลล์ lysis และ
ปล่อย phage ออกจากเซลล์ Temperate phage นอกจากจะมี lytic cycle ได้แล้วยังจะเข้าสู่ขบวนการ lysogenization
คือการทีจีโนม Phage เข้าไปแทรกตัวแฝงอยู่ในโครโมโซมของแบคทีเรียในรูป prophage ได้โดยไม่ทําให้เซลล์
แบคทีเรียแตกสลาย และ prophage จะแบ่งตัวเพิมจํานวนไปพร้อมกับแบคทีเรีย
Transduction
เป็นกระบวนการถ่ายทอดสารพันธุกรรมของแบคทีเรียโดยไวรัส เกิดขึนโดยมีการรวมชินส่วน DNA ของแบคทีเรียเจ้า
บ้านเข้าไปในไวรัส เมือไวรัส infect แบคทีเรียข้างเคียง ทาให้แบคทีเรียทีสองนีมีชินส่วน DNA ของแบคทีเรียแรกเข้าไป
1. Generalized transduction
เกิดในระหว่าง lytic cycle ในวงจรชีวิตไวรัส ชินส่วนของ DNA แบคทีเรียได้
เข้าไปอยู่ในอนุภาคไวรัสระหว่างการประกอบอนุภาคไวรัสใหม่ เมืออนุภาค
ไวรัสตัวนี infect แบคทีเรียตัวใหม่ ซึงไม่ทําให้เกิด lytic cycle ตัว DNA ทีเข้า
ไปนีอาจจะรวมเข้าเป็นส่วนหนึงของจีโนมของแบคทีเรียหรือไม่ก็ได้
2. Specialized transduction
เกิดเนืองจากความผิดพลาดใน lysogenic cycle ในวงจรชีวิตของไวรัส เมือ
prophage ถูกกระตุ้นให้เข้าสู่ lytic cycle โดยแยกตัวออกจากจีโนมของ
แบคทีเรีย แต่การแยกตัวนันไม่ถูกต้อง โดยมีการนําชินส่วนของจีโนม
แบคทีเรียออกมาด้วย เมือเข้าเข้าสู่ lytic cycle อนุภาคไวรัสใหม่ก็จะมี DNA
ของแบคทีเรีย เมือไวรัส infect แบคทีเรียข้างเคียงก็จะทําให้แบคทีเรียได้รับ
DNA ใหม่เข้าไป
การนําไปใช้ในงานพันธุวิศวกรรม
1. ใช้ phage เป็น vector ในการนํายีนเข้าสู่เซลล์แบคทีเรีย
ในการทํา DNA cloning เช่นทําโดยวิธี transduction โดยการนํา DNA สายผสมใส่เข้าในหัวของ phage ก่อน เรียกว่า
in vitro packaging แล้วนําอนุภาคไวรัสนีไปติดเชือแบคทีเรียต่อไป
lambda phage สามารถใช้เป็น vector นํายีนขนาด 15 kb ถ้าต้องการนํายีนขนาดใหญ่กว่าต้องใช้ cosmid
เป็นยีนพาหะโดย cosmid มีลักษณะคล้าย plasmid สร้างจากการนํา DNA ของ phage lambda ส่วนปลายทีเรียกว่า
cos (มาจาก cohesive end) ทีอยู่ทีปลายทังสองข้างมาต่อกับ DNA plasmid ของแบคทีเรียส่วน Ori (origin of
replication) พร้อมทังมีส่วนทีควบคุมการดือยาและส่วนทีจะถูกตัดด้วย restriction endonuclease อยู่ด้วย
cosmid สามารถนํามาใส่ในหัวของ phage lambda ได้ และสามารถนํายีนขนาด 35-45 kb ซึงใหญ่กว่ายีนที plasmid
จะนําได้ (ขนาด 100-5,000 เบส) ในขณะทีปกติ transducing phage จะนํายีนขนาด 5-20 kb เท่านัน
2. การนําไปใช้ทํา phage typing
ใช้ในการจําแนกสายพันธุ์ของแบคทีเรียโดยอาศัยความจําเพาะของชุด (panel) ของ phage กับแบคทีเรีย
ใช้สืบสวนหาต้นตอการระบาดของโรค
3. การ transduction
ขบวนการทีใช้ phage พายีนของแบคทีเรียผู้ให้ ไปยังแบคทีเรียผู้รับ Phage ทีทาให้เกิด transduction ได้
เรียกว่า transducing phage
- 21. 4. การเกิด phage conversion
เป็นขบวนการที lysogenic phage ทําให้แบคทีเรียมีคุณสมบัติแตกต่างไปจากเดิมทําให้ somatic antigen
ของ Salmonella เปลียนแปลงทําให้เกิดการสร้าง diptheria toxin, botulinum toxin, erythrogenic toxin โดยการที
lysogenic phage นํายีนสาหรับสร้าง toxin เข้าสู่แบคทีเรีย
GENE CLONING
เรียกอีกอย่าง พันธุวิศวกรรม (genetic engineering) , เทคโนโลยีดีเอ็นเอสายผสม (Recombinant DNA Technology)
พันธุวิศวกรรม หมายถึง กระบวนการเปลียนแปลงหรือดัดแปลงสารพันธุกรรมของสิงมีชีวิต โดยการ
ถ่ายทอดยีนทีต้องการจากสิงมีชีวิตหนึง เข้าสู่อีกสิงมีชีวิตหนึง เพือสร้างสิงมีชีวิตชนิดใหม่ทีมี ลักษณะตามต้องการการ
ตัดต่อดีเอ็นเอทีมาจากหลายต้นตอในหลอดทดลอง โดยอาศัยเอนไซม์ตัดจําเพาะ แล้ว เชือมต่อชินดีเอ็นเอเข้าไปใน
พาหะ ทําให้ได้ดีเอ็นเอโมเลกุลใหม่ทีใส่เข้าไปกับดีเอ็นเอพาหะ เรียกว่า ดีเอ็นเอสายผสม (Recombinant DNA )
ขันตอนการทําพันธุวิศวกรรมอาจเรียกว่า gene cloning คือ การเพิมปริมาณกลุ่มของเซลล์ทีมีลักษณะทาง พันธุกรรม
เหมือนกันซึงมียีนทีต้องการอยู่ในเซลล์ให้ได้จํานวนยีนและจํานวนเซลล์ในปริมาณมากพอทีจะนําไปใช้ ประโยชน์ต่อไป
ขันตอนการทํา Gene cloning
1. การเตรียมชินส่วนของ DNA ทีต้องการ (Preparing DNA)
2. การใช้เอนไซม์ตัดจําเพาะในการโคลนยีน (Restriction enzyme for gene cloning)
3. การเลือกใช้พาหะหรือเวคเตอร์ (Vector)
4. การถ่ายไฮบริดเวคเตอร์เข้าสู่เซลล์เจ้าบ้าน เพือทํา Gene cloning (Transformation)
5. การตรวจสอบ clone ทีต้องการ (Screening hybrid vector)
6. การประยุกต์ใช้เทคนิคพันธุวิศวกรรม (Application)
I. การเตรียมชินส่วนของ DNA ทีต้องการ
การเตรียมชินส่วนยีนหรือดีเอ็นเอทีน่าสนใจมีหลายวิธีดังนี
1. การสกัดจากเซลล์หรือเนือเยือของสิงมีชีวิตทีต้องการศึกษา ซึงเป็นดีเอ็นเอทังหมดในจีโนมของสิงมีชีวิต ชนิดนัน
เรียกว่า genomic DNA
2. การเตรียมดีเอ็นเอจาก mRNA
– โดยใช้เอนไซม์ reverse transcriptase
– จะได้ดีเอ็นเอทีสังเคราะห์ขึนหรือลอกรหัสจาก mRNA เรียกว่า complementary DNA (cDNA)
3. การสังเคราะห์ดีเอ็นเอโดยวิธีทางเคมี
– สามารถสังเคราะห์ oligonucleotide ให้มีลําดับเบสทีต้องการโดยเครืองสังเคราะห์อัตโนมัติ
– มี 2 วิธีทีใช้กันอย่างกว้างขวางคือ วิธี phosphate triester และวิธี phosphite triester
II. เอนไซม์ตัดจําเพาะในการโคลนยีน
ในการโคลนยีนต้องใช้เอมไซม์ตัดจําเพาะในการตัดสายดีเอ็นเอทีต้องการ จากนันนําไปเชือมต่อกับพาหะ (Vector) แล้ว
จึงพาหะถ่ายเข้าสู่เซลล์เจ้าบ้าน (Host cell)
- 22. • เอนไซม์ตัดจําเพาะ(Restriction enzyme or restriction endonuclease)
• แบบที 2 (type II) เป็นเอนไซม์ทีใช้กันมากในการตัดต่อยีน
• ประกอบด้วยโพลีเพพไทด์เพียงชนิดเดียว
• มีคุณสมบัติเป็นเอนไซม์ตัดจําเพาะเพียงอย่างเดียว ส่วนการเติมหมู่เมธิลอาศัยเอนไซม์อีกชนิดหนึง
• การตัดดีเอ็นเอจะเกิดทีตําแหน่งจําเพาะในบริเวณจดจํา(recognition site)หรือจุดใกล้กับบริเวณจดจํา ทําให้ได้ชิน
ขนาดดีเอ็นเอทีมีขนาดแน่นอน
ปัจจุบันเอนไซม์ตัดจําเพาะทีสกัดได้จากแบคทีเรียชนิดต่างๆ มีจํานวนมากว่า 400 ชนิด ซึงการเรียกชือใช้
ระบบอักษรภาษาอังกฤษ 3 ตัว และพิมพ์ตัวเอน โดยอักษรตัวที1 คือ อักษรตัวแรกของชือ Genus ใช้เป็นตัวพิมพ์ใหญ่
อักษรตัวที 2 และ 3 เป็นอักษร 2 ตัวแรกของชือ Species ใช้ตัวพิมพ์เล็ก ถ้ามีรหัสของสายพันธุ์ก็ให้ใส่หลังอักษร 3 ตัว
แรก และสุดท้ายเป็นเลขโรมันซึงบอกถึงลําดับของเอนไซม์ทีสกัดได้จากแบคทีเรีย เช่น
EcoRI สกัดมาจาก Escherichia coli RY13 แยกได้เป็นชนิดแรก
HindIII สกัดมาจาก Haemophilus influenzae แยกได้เป็นชนิดที 3
III. การเลือกใช้พาหะ (Vector) และการทํา Gene cloning
Vector คือ ดีเอ็นเอพาหะทีใช้ในการเพิมปริมาณชินดีเอ็นทีต้องการให้ได้ปริมาณมากๆ โดยนําชินดีเอ็นเอที
ต้องการดังกล่าวมาเชือมต่อเข้าไปกับ DNA vector ได้ ดีเอ็นเอสายผสม (recombinant DNA; rDNA)
จากนันจึงนํา rDNA ถ่ายเข้าสู่เซลล์เจ้าบ้าน (Host cell) โดยวิธีการ Transformation เพือเพิมปริมาณดีเอ็นเอ
ทีต้องการ แล้วทําการคัดเลือกเซลล์ทีต้องการต่อไป
ปัจจุบันมีเวคเตอร์ต่างๆ มากมายทีใช้สาหรับ Gene cloning ซึงการเลือกใช้เวคเตอร์นันขึนอยู่กับวัตถุประสงค์
ของการใช้งาน เช่นถ้าต้องการโคลนยีนในแบคทีเรีย อาจเลือกใช้เวคเตอร์ทีเป็น พลาสมิด (Plasmid) ฟาจ (Phage) และ
คอสมิด (Cosmid)
คุณสมบัติของดีเอ็นเอพาหะ(vector) มีดังนี
- มีส่วนของดีเอ็นเอทีเป็นจุดเริมในการจําลองตัว (origin of replication,Ori)
- สามารถเพิมจํานวนตัวเองได้พร้อมๆกับดีเอ็นเอทีใส่เข้าไป
- มียีนเครืองหมาย(marker gene) สาหรับใช้ในการคัดเลือก เช่น ยีนทีดือยาปฏิชีวนะ ยีนทีเกียวกับการสร้างหรือสลาย
กรดอะมิโนและนําตาลบางชนิด
- มีบริเวณจดจําของเอนไซม์ชนิดใดชนิดหนึงหรือหลายชนิดเพียง ตําแหน่งเดียวในโมเลกุลของ
ดีเอ็นเอพาหะ(unique or polylinker site)
- ไม่อยู่ในบริเวณทีเป็น replicon
- ควรอยู่ในยีนทีเป็น marker เช่น lac Z gene (b-galactosidase)
- เมือนํา foreign DNA มาเชือมจะทําให้เกิดการทํางานของยีนดังกล่าวบกพร่อง เรียกว่า marker inactivation
- 23. IV. การเชือมชินดีเอ็นเอเข้ากับเวคเตอร์ หรือ Gene cloning
Gene cloning หมายถึง การนํายีนทีต้องการหรือยีนทีสนใจมาเพิมปริมาณให้ได้เพียงพอเพือใช้ในการศึกษา อาทิ การ
แสดงออกของยีน การผลิตโปรตีนหรือเอนไซม์ การถ่ายยีน การหาลําดับเบสของยีน เป็นต้น ซึงหลักการทํา Gene
cloning ก็คือ หลังจากได้ rDNA (recombinant DNA) แล้วก็นําไปถ่ายลงในเซลล์เจ้าบ้านเพือเพิมปริมาณ ซึง การทํา
rDNA นันสามารถทําได้หลายวิธี ดังนี
1. การสร้าง DNA library
คือ การนํา Genomic DNA ของสิงมีชีวิตมาตัดด้วยเอนไซม์ตัดจําเพาะ แล้วนํามาเชือมต่อเข้ากับเวคเตอร์ซึงถูกตัดด้วย
เอนไซม์ตัดจําเพาะชนิดเดียวกัน จากนันจึงนําไปถ่ายในเซลล์เจ้าบ้านเพือขยายปริมาณชินดีเอ็นเอ แล้วเก็บเซลล์เจ้า
บ้าน
2. การเชือมต่อชินดีเอ็นเอทีต้องการเข้ากับเวคเตอร์
สามารถแบ่งได้เป็น 2 วิธี ดังนี
- การเชือมต่อดีเอ็นเอปลายเหนียว
- การเชือมต่อดีเอ็นเอปลายทู่
3.การเพิมปริมาณ DNA เป้ าหมาย หรือยีนในเซลล์ผู้รับ
หลังจากทีตัดต่อและถ่ายเข้าไปแล้ว ถ้าเป็นในเซลล์แบคทีเรีย สามารถทําได้โดยเลียงแบคทีเรียในอาหารเพาะเลียงเชือ
เพือให้ได้หลายโคโลนี และนําแบคทีเรียเหล่านันไปสกัดเอายีนทีต้องการหรือให้แบคทีเรียสร้างสารทีเราต้องการ
ให้ bacteria สร้างสารทีเราต้องการ เพิมจานวนbacteria ทีมี recombinant gene อยู่
- 24. V. การตรวจสอบ clone ทีต้องการ
หลังจากใส่ชินดีเอ็นทีต้องการลงไปในเวคเตอร์ แล้วนําเวคเตอร์ดังกล่าวมา Transformation เข้าสู่ Host cell ในขันตอน
นีจําเป็นมีวิธีการตรวจหาโคลนทีต้องการจากประชากรทังหมดของเซลล์เจ้าบ้าน แล้วคัดเลือกโคลนทีต้องการนัน
ออกมา เพือเพิมจํานวนต่อไป
การคัดเลือกโดยอาศัยลักษณะ (phenotype)
เป็นการอาศัยการแสดงออกของยีนเครืองหมาย (marker gene) และมีการสร้างโปรตีนออกมา
เช่น การสร้างเอนไซม์บางชนิดทีทําให้เกิด clear zone ในอาหารเลียงเชือทีมี substrate (IPTG) อยู่
หรือพบลักษณะการดือยาในเซลล์ผู้รับทีได้รับมาจากพลาสมิด
การคัดเลือกโดย DNA hybridization
อาศัยการจับคู่กันของสายดีเอ็นเอทีเป็นตัวติดตาม (probe) กับดีเอ็นเอเป้าหมายทีต้องการตรวจหาทีมีลําดับเบสคู่สม
กัน บน membraneโดย probe จะติดฉลากด้วยสารไอโซโทปหรือเอนไซม์
การคัดเลือกโดยวิธีทางอิมมูโนวิทยา
ทําได้เมือโคลนทีต้องการแสดงออกได้ โดยผลิตโปรตีน แต่โปรตีนไม่แสดงลักษณะหรือ phenotypeทีชัดเจน เป็นการ
ตรวจสอบโปรตีนทีเซลล์ผลิตขึนโดยใช้แอนติบอดีทีจําเพาะกับโปรตีนทีต้องการนัน
การประยุกต์ใช้พันธุวิศวกรรม
การวินิจฉัยโรค
ปัจจุบันมีการนําเอาเทคโนโลยีของDNAมาใช้ในการวินิจฉัยโรคทีเกิดจากการติดเชือต่างๆ เช่น เชือไวรัส
โดยการใช้เทคนิคPCR เพือตรวจสอบว่ามีจีโมนของไวรัสอยู่ในสิงมีชีวิตนันหรือไม่ ซึงเป็นเทคนิคทีมีความไวสูงและ
สามารถตรวจพบได้โดยมีตัวอย่างเพียงเล็กน้อย เทคนิคนีได้นํามาใช้ในการตรวจวิเคราะห์การติดเชือ HIVเป็นต้น
การบําบัดด้วยยีน
จากความรู้เกียวกับความผิดปกติต่างๆในคนทีเกิดความบกพร่องของยีน หากสามารถใส่ยีนทีปกติเข้าไปใน
เซลล์ร่างกาย หรือเนือเยือทีแสดงอาการผิดปกติ แล้วทําให้ยีนนันแสดงออกเมือมีสารโปรตีนทีปกติในบริเวณดังกล่าว
จึงอาจเป็นแนวทางหนึงทีจะช่วยทําให้บําบัดอาการบกพร่องทีเกิดขึนได้ ในปัจจุบันเทคนิคหนึงทีใช้ในการถ่ายยีนปกติ
เพือใช้ในการทํายีนบําบัด คือการใช้ไวรัสชนิดหนึงเป็นตัวนํายีนทีต้องการถ่ายเข้าสู่เซลล์คน ซึงยีนของไวรัสทีเป็น
อันตรายต่อคนจะถูกตัดทิง แล้วใส่ยีนของคนทีต้องการเข้าไปแทนที ไวรัสทีสร้างขึนใหม่นีจะมียีนทีต้องการแทรกอยู่
และจะมีความสามารถในการแทรกจีโนมของตัวมันเข้าสู่โครโมโซมคนได้ แต่ไม่สามารถจําลองตัวเองเองเพิมจํานวนได้
เนืองจากยีนทีทําหน้าทีดังกล่าวทีมีอยู่เดิมในไวรัสได้ถูกตัดทิงไป
การสร้างผลิตภัณฑ์ทางเภสัชกรรม
การผลิตฮอร์โมนอินซูลิน เป็นตัวอย่างแรกทีทีนําเทคนิคทางDNAมาใช้ในการผลิตสารทีใช้เชิงเภสัชกรรมเพือรักษา
โรคเบาหวาน ผู้ป่วยโรคเบาหวานจําเป็นต้องได้รับอินซูลิน เพือควบคุมระดับนําตาลจากการตัดและต่อDNAให้มียีนที
สร้างอินซูลิน แล้วใส่เข้าไปในเซลล์แบคทีเรีย เพือให้เกิดการแสดงออกและสร้างพอลิเพปไทด์ทีต้องการ จากนันจึงนํา
เซลล์ไปเพือเพิมจํานวนยีนทีสร้างสายพอลิเพปไทด์ดังกล่าว และผลิตอินซูลินทีทํางานได้
- 25. การผลิตฮอร์โมนอินซูลินโดยใช้เทคนิคพันธุวิศวกรรม
มีขันตอนโดยสรุป ดังนี
1. การตัดชินส่วน DNA ของคนทีมียีนควบคุมการสังเคราะห์
ฮอร์โมนอินซูลิน ต่อเข้ากับ DNA ของแบคทีเรีย จะได้ DNA
โมเลกุลใหม่ เรียกว่า DNA สายผสม
2. นํา DNA สายผสมใส่เข้าไปในเซลล์ของแบคทีเรีย ทีไม่
สามารถสังเคราะห์ฮอร์โมนอินซูลิน ให้กลายเป็นแบคทีเรีย
ชนิดใหม่ทีสามารถสังเคราะห์ฮอร์โมนอินซูลินได้
3. คัดเลือกแบคทีเรียชนิดใหม่ทีสามารถสังเคราะห์ฮอร์โมน
อินซูลินได้และนําไปเพาะเลียงให้มีเซลล์จํานวนมากเพียงพอ
แล้ว จึงสกัดฮอร์โมนอินซูลินไปใช้ประโยชน์ต่อไป
- 26. พืนฐานเบืองต้น (biology of cell)
THE CELL
Cell
Protoplasm Cell membrane
Cytoplasm Nucleus
Organelle Inclusion
Membrane bound Non-membrane bound Exogenous Endogenous
Cell membrane
ประกอบด้วย Lipid = phospholipids(phosphoglyceride,sphingolipid), steroid(cholesterol)
Protein = extrinsic(peripheral) และ intrinsic(integral)
Carbohydrate = glycocalyx(cell coat) , glycolipid , glycoprotein
เรียงตัวแบบ Fluid-Mosaic model
Protoplasm
Cytoplasm
Ectoplasm มี actin filament มาก
Endoplasm มี membrane bound organelle มาก จึงเรียกร่างแหทีพบใน endoplasmว่า Endoplasmic reticulum
Organelle
1. Endoplasmic reticulum คือ membrane bound organelle ทีเป็นร่างแห่ของ vesicle , เยือหุ้ม ER เชือมต่อกับ
cell membrane , Golgi , nuclear membrane โดยหุ้มโพรงทีเป็นท่อ เรียก lumen หรือ ER cisternal space
RER = ER ทีมี ribosome มาเกาะอยู่ หน้าที คือ การสังเคราะห์โปรตีน(Secretary protein)ของไรโบโซม
ทีเกาะอยู่ โดยมี Golgi body เป็นตัวสะสม หรือทําให้มีขนาดพอเหมาะ ทีส่งออกนอกเซลล์ ลําเลียงสาร ซึงได้แก่
- 27. โปรตีนทีสร้างได้ และสารอืนๆ เช่น ลิพิดชนิดต่างๆ. ในเซลล์ทีเกิดใหม่ พบว่ามี RER มากกว่า SER แต่เมือเซลล์มี
อายุมากขึน พบว่า SER มากกว่า RER เชือกันว่า RER จะเปลียนเป็น SER เมือเซลล์มีอายุมากขึน พบในตับอ่อน
ลําไส้เล็ก ต่อมใต้สมอง
SER = ER ทีไม่มี ribosome มาเกาะ , พบ Mitocodria ทีมี tubular cistae
หน้าที 1. Lipid synthesis = มี Enz. ทีสําคัญในการสังเคราะห์ Lipid รวมถึง phospholipid และ Steroid
2. Detoxification of drug and poison = Enz. ใน SER ทําให้สารพิษละลายนําได้เพือขับออกทางไต
3. Calcium turnover = ควบคุมระดับ Ca2+
ในเซลล์ โดยเฉาะการหดตัวของกล้ามเนือ
4. Carbohydrate metabolism ในเซลล์ตับ SER เปลียน glucose phosphate ทีได้มาจาก glycogen ให้เป็น
glucose เพือขับออกจากเซลล์ตับ
พบใน เซลล์ทีต่อมหมวกไต , Leydig’s cell ในอัณฑะ , เซลล์คอร์ปัสลูเทียมในรังไข่ และในเซลล์ของตับ
2. Ribosome คือ non-membrane bound organelle เกิดจาก ribosomal RNA (rRNA) จับกับ ribosomal protein
เป็น ribosome subunit ขนาดเล็ก(30s,40s,50s,60s โดย 30s+50s = 70s และ 40s+60s = 80s). โดยทําหน้าที
สังเคราะห์โปรตีนในกระบวนการ Translation. Ribosome มี 3 form ได้แก่ free form , polysome , rER
3. Golgi apparatus เกิดจาก Golgi cisterna หลายอันซ้อนทับกัน ซึงทางด้านโค้งเรียก cis face และทางด้านเว้า
เรียก trans face.
หน้าที 1. Protein maturation โปรตีนทีสร้างจาก RER เป็น immature ซึงจะทําให้เป็น mature ที Golgi
2. Protein sorting การคัดแยกโปรตีนชนิดเดียวกันไว้ในถุงเดียวกัน
3. Protein condensation การทีโปรตีนอืนไม่
4. Membrane equilibrium เกิดตอนกระบวนการ exocytosis ทําให้ membrane เพิมขึนและ membrane ที
เพิมขึนนีก็จะถูกนํากลับไปให้ Golgi ด้วย endocytosis
4. Lysosome คือ membrane bound organelle ทีเป็น digestive organelle ทําหน้าทีย่อยโดยใช้ Enz. (Hydrolase)
ที active ในกรด (pH 5) ซึงควบคุมความเป็นกรดโดย Proton pump ทีอยู่บน membrane , ภายในบรรจุ Hydrolase
มากกว่า 40 ชนิด (ทีสําคัญคือ acid phosphate) , พบมากใน phagocyte
5. Endosome คือ membrane bound organelle ทีได้จากกระบวนการ Endocytosis
6. Proteasome คือ non-membrane bound organelle ทีประกอบด้วย protein complex(core protein + regulatory
protein) โดยมีคุณสมบัติเป็น ATPase ทําหน้าที ย่อยโปรตีนในcytosol(proteolysis) และ subunitบางตัวทําหน้าทีจับ
กับ ubiquitin ทีตําแหน่ง ubiquitin recognizing site ของ regulatory protein , ubiquitin คือ โปรตีนขนาดเล็ก (76
amino acid)
7. Peroxisome มีลักษณะคล้าย Lysosome ภายในบรรจุ oxidizing enz. กว่า 40 ชนิด รวมทัง D-aminooxidase
และ catalase หน้าทีคือ detoxify toxic substance และ neutralizes free radical โดย oxidase
8. Mitocondria คือ double membrane bound organelle โดยแบ่งเป็น inner และ outer ซึง